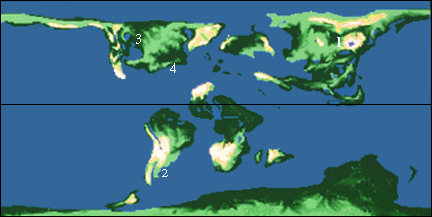
Locations are: 1, Argentina; 2, Two Medicine Formation, Montana; 3, Judith River beds, Montana and Alberta; 4, Hell Creek Formation, Wyoming and Montana; 5, New Jersey
BACK TO DINOSAURS 2001 HOME PAGE
Locations are: 1, Argentina; 2, Two Medicine Formation, Montana; 3, Judith River beds, Montana and Alberta; 4, Hell Creek Formation, Wyoming and Montana; 5, New Jersey
Saltosaurus was a small sauropod characterized by a diplodocid-type head (teeth in anterior of mouth only), but with a body covered with small bony plates imbedded in the skin. The later is the shared derived character for the group of sauropods called the Titanosauridae.
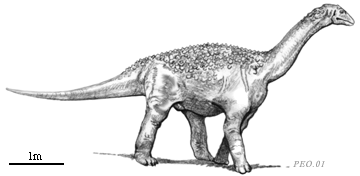
Saltosaurus

Armor of Saltosaurus, after xxxx. |
When just the bony plates were found they were originally ascribed
to that of ankylosaurs, which the plates resemble.
For this course we regard the Titanosauridae as one other the three groups of sauropods in an unresolved polytomy. In fact, the Titanosauridae are probably a group within the diplodocids. While abundant in South America, members of the Titanosauridae such as Alamosaurus, spread into North America and seem to have lasted until the end of the Cretaceous. |
| Recently (1998) superbly preserved eggs, apparently of sauropod origin
were discovered in Argentina some with preserved embryos inside.
The embryos have skin with the embedded bony plates The overall shape of the eggs is spherical, very much like the Chinese eggs attributed to sauropods. For more information see: http://www.canoe.com/CNEWSScienceArchive/ 981117_dinosaurs.html. |

Titanosaurid eggs, based on xxxx. |
Carnotaurus

 |
Carnotaurus is a large theropod know from skeletal material that includes detailed impressions of skin, including the face. The skull is unusually short for a theropod and its sports two horns over the eyes. The face was covered with large to small scales as shown on left. The name Carnotaurus means meat-eating bull. |

Based on xxxx.
The skull and skeleton of Carnotaurus shows similarities to Allosaurus
as well as certatosaurians. We group it with the latter here although I
strongly suspect it should be with the Carnosauria.
| Without benefit of a formal bachelor degree from a university, he came into paleontology as an amateur collector, after a tour of duty in the Vietnam War, Horner became a vertebrate paleontological preparator for the former Museum of Natural History at Princeton University, under the then Director and Curator, Donald Baird. There he was able to have free access to an excellent Library and during the summers spend much time collecting in his beloved Montana, especially dinosaurian remains in the Two Medicine Formation. |  |
| He did other collecting as well, however. Above right we see Horner collecting a partial prosauropod dinosaur with Donald Baird that I had found in 1975. This skeleton, found at Wasson Bluff near Parrsboro Nova Scotia, was the first Early Jurassic dinosaur and the first prosauropod dinosaur found in Canada. | |
|
Horner (on left) always kept in touch with other amateurs, and in 1978, he was shown some small bones by owners of a rock shop in Choteau, Montana, Marion and John Brandvold. Horner recognized these to bones of a baby hadrosaur. They showed Horner where these came from and Horner began to find enormous numbers of dinosaur egg fragments, and eventually nests of eggs as well as many dinosaur bones. Many of the eggs, nests, babies, and adults turned out to be remains of a new hadrosaur, that Horner named Maiasaura peeblesorum, or "good mother lizard". Horner prepared much of the material at Princeton, where he gained fame, eventually accepting a position at the Museum of the Rockies in Montana. He became Curator of Paleontology xxxx and received an Honorary Doctorate of Science in 1986 from the University of Montana. Horner also was technical advisor for the movies Jurassic Park and The Lost World, directed by Steven Spielberg. |
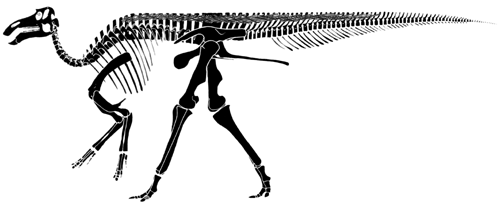
Modified from Carpenter in Horner and Gorman (1988)
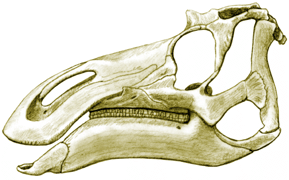
Skull of Maiasaura peeblesorum; based on Horner, 1988. |
The skull of Maiasaura peeblesorum is also fairly typically
non-crested hadrosaurian.
It has the inflated narial (nostril) area typical of most hadrosaurs as well, but one not quite as large as is typical. |
| But, of course its the eggs and babies that Maiasaura
is really famous for.
Below left is an example of a nest of eggs and below right a cutaway view of what the fetus might look like within the egg, just before hatching. |
|

Based on Horner, 1982. |

Based on Horner, xxxx. |
A map of the distribution of nests suggests that they were spaced about one Maiasaura apart and that they nested in large numbers.

Based on Horner (1987)
Horner has shown that there is plant matter preserved within the fossil nests and is might have been brought for protection and warmth, or it might have been food for the babies.
That the babies at least returned to the nest for quite a while is shown by the variety of sizes of juveniles that have been found within the nests. Below is a reconstruction of a hatching Maiasaura, based on xxxx.

Horner suggests that these sorts of bone beds were created by the mass-death of huge herds of hadrosaurs, caused by volcanic ash falls.
Varricchio and Horner (1993) estimate that at least xxxxxxx individuals are in the largest of these bone beds, and that these probably came from a single vast herd. Such estimates are made by counting the number of only one kind of bone (e. g, right tibia) within a given volume and then multiplying this number times an estimate of the total volume of the bed.
Such a herd, migrating across the landscape undoubtedly had a massive
effect on the plant communities.
Troodon is the type genus for the Troodontia. While incomplete skeletal material of adult Troodon are known (including the characteristic teeth) from the Two Medicine Formation, Horner has found nests with eggs and babies, of these dinosaurs as well.

 |
Henry Fairfield Osborn was first hired at the American Museum of Natural History in 1891 on the suggestion of the president of Columbia University. As pointed out by Preston (1986), at that time the museum had not a single dinosaur skeleton. In 1897, Osborn hired Barnum Brown (left) who had made himself known through part time work on collecting trips. Brown eventually collected many of the most spectacular skeletons in the American Museum collection including the magnificent Apatosaurus skeleton from the Morrison and Tyrannosaurus rex skeletons from the Hell Creek. |
Saurischia
Without a doubt, the most famous Hell Creek dinosaur is the other arctometatarsalian tyrannosaur, Tyrannosaurus rex.

Redrawn from Paul (198x).
Barnum Brown collected several excellent specimens of Tyrannosaurus
rex for the American Museum of Natural History. Including the spectacular
skeleton on exhibit as well as the huge skull. The name means "king of
the tyrant lizards" and apart possibly from Giganotosaurus, Tyrannosaurus
rex is the largest known carnivore to have lived. Another skeleton
of T. rex collected by Brown was given to the Carnegie Museum of
Pittsburgh PA, when it was thought New York might be bombed during WW II
(Preston, 1986).
 |
The forelimbs of Tyrannosaurus rex are proportionally very small, even for a tyrannosaur (left). However, as pointed out by Bakker (1986) they were absolutely large and quite powerful. As previously discussed, the reduction of the size of the forelimbs and the reduction of the manus to two digits is the shared derived character for the Tyrannosauria. |
| The foot of Tyrannosaurus rex has the characteristic arctometatarsalian proximally pinched metatarsal III. (right). |  |
Exactly what the function of the relatively minute hands of T. rex was, remains controversial. They are too short to reach the head. One idea is that the arms were used to prevent the dinosaur from slipping forward when in trying to get up off the ground. Another, is that they acted as braces against prey, while ripping out large chunks of flesh..

T. rex rising from a nap. Redrawn from xxxx (198x).
No one argues that T. rex was not a meat eater. However, Horner
(xxxx) has hypothesized that it was a scavenger, eating dead carcasses,
and that it was too slow to capture its own prey. Farlow et al. (1995)
have even gone so far as to argue that T. rex was so massive that
if it ran and tripped it would be fatally injured by the fall! Personally,
I find these arguments unconvincing, although it must be stressed that
it is very hard to make behavior arguments for extinct forms when there
is not even a remote living analog.
| There is no doubt, however, that the skull of T. rex was a massive meat-eating machine (right). The over all structure of the skull, like that of other theropods, was strong in tension (while pulling). Indeed, there parts of the skull was unusually flexible - an area lower jaws, allowing the jaw to flex while ingesting particularly bulky prey or bits of prey. | 
Cast of AMNH 5027, collected by Barnum Brown in 1908, on display at the the North Carolina Museum of Natural History. |
 |
The teeth of Tyrannosaurus rex average about xx cm and have a design particularly well suited to ripping out massive chunks of flesh. As described by Paul (1988) each premaxillary tooth is somewhat "D"-shaped in cross-section (left). As a bite proceeded the blunt side of the tooth would be relatively ineffective at cutting, but would tend to push the tooth posteriorly as the tooth was pushed into the flesh. The rear cutting edge would thus be forced backward and inward at the same time. This action would have automatically sliced out flesh in the bite stroke. |
Brown collected a large number of ornithischians from the Hell Creek beds, mostly Triceratops. However other dinosaurs included hadrosaurs and ankylosaurs. The remains of Ankylosaurus comprise the most common ankylosaur in the Hell Creek Formation. Obviously it is a member of the Ankylosauridae and displays the characteristic shared derived characters of the group.
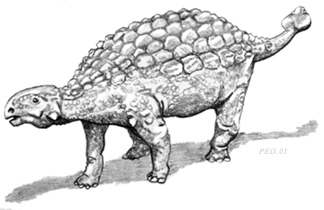
Reconstruction of Ankylosaurus
 |
The skull of Ankylosaurus is expanded transversely (right),
unlike that of nodosaurids. Missing is the central forehead plate of nodosaurids
as well (retention of a primitive character).
The tail has the main shared derived character of the Ankylosauridae, a massive club at the end of a stiffened tail (right). Note that the distal tail vertebrae have interlocking processes that stiffened the tail to support swings of the massive club. |
 |
Ankylosaurus is not as well known as its relative Euoplocephalus,
and therefore additional comments on the Ankylosauridae will be dealt with
under the latter genus, below, when we consider the Judith River beds.
Anatotitan
Anatotitan, or as it is perhaps better known by is younger synonym Anatosaurus, is the quintessential "duck billed dinosaur". Its name means "titan duck" and images of its skeletons and reconstructions have appeared in popular books since Brown's beautiful set of skeletons was mounted.

Based on xxxxxxxxx
Anatotitan at the Tyrell Museum |
The skull of Anatotitan is unusual because in lateral view (left) it is low and, sleek. A dorsal view (right), however, shows the broad expansion as the anterior of the skull that supported the duck-like bill. |  |
| Of course, unlike birds, Anatotitan has a typically hadrosaurine dental battery. This dental battery consists of teeth set in a regular alternating pattern making a dense stack in the jaw. Growing teeth at the base of the dental battery displace the entire apparatus through the jaw. Wear on vegetable matter and the opposing teeth wear the entire batter down and n the process create a broad rasp-like surface comprised of obliquely work teeth, The surface is kept ever fresh by growth from the base of the battery and the wear. Weishample (1984) has shown that the lower jaws moved against the upper jaws in a circular motion, with the two lower jaws moving in mirror image motions against the upper jaws. |  |
Edmontosaurus

Edmontosaurus, based on xxxxx.
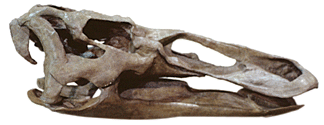
| Edmontosaurus is a very well known non-crested hadrosaur. It was originally described from the Judith River beds, in Alberta - Edmonton being the province's capital. Nonetheless, perhaps the most famous specimen comes from the Hell Creek Formation of Wyoming. It was collected by the famous team of professional collectors the Sternbergs. | 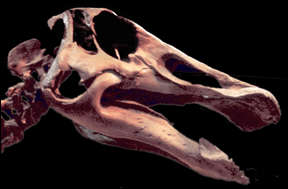
Skull of Edmontosaurus from the Judith River beds (below) on display at the North Carolina Museum of Natural History. |
The Sternberg team consisted of Charles Sternberg and his three sons/ As described by Preston (1986), the Sternbergs were on a collecting trip in Hell Creek beds for a Triceratops skull for the British Museum of Natural History in London. Triceratops is the most common dinosaur in the Hell Creek. One of Charles Sternberg sons, George found a hadrosaur skeleton, and in the process of excavation the elder Sternberg notice that much of the skeleton was covered in skin impressions preserved in the sandstone encasing the Edmontosaurus. This specimen was purchased by the American Museum of Natural History where it gained great fame as "the dinosaur mummy". It is one of the most remarkable specimens on display at the museum.
.
 |
Until recently it was quite common for hadrosaurs to be thought of a semiaquatic. Some evidence for this was supposed to be webbing visible on the Edmontosaurus mummy's hands (left). Bakker (xxxx), however, has pointed out that the supposed webbing is more like loose skin left by decay of thick pads. Bakker also points out that the front feet are minute compared of the hind feet and would have provided minimum propulsive force in swimming. Bakker also argues that although hadrosaur tails were dorsoventrally deep the transverse spines on the anterior caudal vertebrae were very short compared to an animal that uses its tail to swim, like a crocodile. These transverse processes support that muscles that move the tail from side to side. |
| Footprints described by Lockley and Hunt (1995) show that hadrosaurs, such as Edmontosaurus clearly walked both bipedally and quadrupedally, and on land. |  |
Triceratops
When the first horn cores of Triceratops were discovered in
xxxx and brought to O. C. March, they were proclaimed an extinct bison.
But when additional skulls were uncovered they proved to be of an extraordinary
three horned dinosaur, Marsh named Triceratops. Barnum Brown later
collected many specimens for the American Museum, a wonderful example of
which is presently on display.

Triceratops, based on Dodson (1996).
| Unlike most other neoceratopsians, the frill on the skull of Triceratops is solid, lacking the large fenestrae usually present. There are nubbins of bone around the frill that must have given it a serrated appearance. there is a short, upward pointing horn on the nose and much longer forward pointing horns over the eyes. |  |
 |
There is considerable debate on whether how neoceratopsians ran. Bakker (xxxx; xxxx) has argued that they could gallop, like a rhinoceros. Recall that a gallop is a kind of run in which all four feet are off the ground in part of the step cycle. When this was first proposed it was considered a radical idea. However, subsequently, Webb et al. (1982) has shown that some crocodiles can gallop, so it seems perhaps less surprising. However, Dodson (1996) claims that the front legs were held in a somewhat sprawling position, which would make galloping very difficult. |
| Lockley and Hunt (1995) describe putative neoceratopsian tracks and suggest that their forelimbs were not held in a sprawling position, but Dodson (1996) looking at the same kind of data suggest otherwise. To me, such speculation is premature, because criteria for recognizing neoceratopsian tracks have not been established in the first place. |  |
There are other kinds of neoceratopsians, with varied horns and frills, found with Triceratops, but these will be discussed below.
Triceratops and Tyrannosaurus rex were certainly among
the last of the dinosaurs. Their remains are found within a few meters
of the Cretaceous-Tertiary boundary and there is little reason to believe
they did not make it right to the end.
Two very well preserved members of the Ankylosauria (within the Tyreophora) are known from the Judith River fauna. These are the nodosaurid Edmontonia and the ankylosaurid Euoplocephalus.

Edmontonia, based on xxxx (xxxx).
Edmontonia is a typical nodosaurian with a plate in the middle of the forehead and a slender tapering tail, lacking club. It has, however, prominent shoulder spikes.
Euoplocephalus (below) is a typical ankylosaurid, and the best known genus in the family. Like Ankylosaurus, it has a massive club at the end of a stiffened tail.

Euoplocephalus, based on xxxx (xxxx).
 |
Nodosaurids and ankylosaurids developed elaborate nasal systems, showing some parallels to mammals, but do it is somewhat different ways. On the left are cross-sections through the skulls of Edmontonia and Euoplocephalus. The nodosaurid Edmontonia has a very complex nasal passage with a fully developed hard palate. Euoplocephalus, the ankylosaurid, as a less complex system with more anterior internal nostrils (choanae). Presumably these features developed to allow more vigorous simultaneous chewing and breathing, as in mammals. |
Thescelosaurus
While the ornithopod Hypsilophodon has traditionally been the most famous hypsilophodontid, Thescelosaurus from the Judith River beds is known from much better and more abundant material, including one specimen that may preserve a heart.

Thescelosaurus, based on xxxx (xxxx).
Thescelosaurus resembles Hypsilophodon in most respects, however, it is larger and seems to have a proportionally smaller skull. Based on recent analysis, it may be more primitive than Hypsilophodon, despite its younger age.
In Spring 2000 an article on the front page of the New York Times
announced the discovery and analysis of a heart preserved in the thorax
of a Thescelosaurus skeleton nicknamed "Willow" in the collections
of the North Carolina Museum of Natural History.
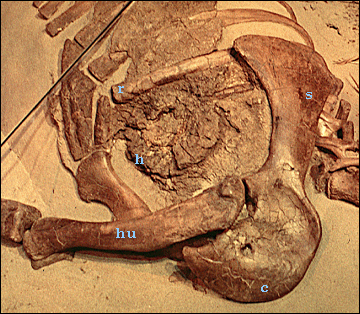 |
The putative heart (left) is preserved as a concretion within the chest
cavity, which shows upon CAT-scanning, what appears to be a four chambered
heart, according to Fisher et al. (2000). According to the latter, the
fossilized heart is four chambered, with fully separated ventricles. If
correctly interpreted, this would mean that ornithischians had a bird-
(or mammal-) like heart with complete separation of the blood pumped through
the body (deoxygenated) and blood pumped thorough the lungs (oxygenated)
as seen on right.
Abbreviations are: c, right coracoid; h, heart; hu, humerus; r, rib; s, scapula. |
| Such a separation is only partial in crocodilians, the living sister group of dinosaurs and birds, and there is only a three chambered heart in living non-archosaurian sauropsids. The three chambered heart mixes oxygenated blood and oxygen depleted blood which makes sustained activity difficult. Because a four chambered heart is present in the avian theropods, if one was present in Thescelosaurus, the most parsimonious hypothesis is that it was primitive for dinosaurs in general, adding more evidence to the argument that dinosaurs had high activity levels and were metabolically similar to birds and mammals. |  |
Parasaurolophus
The Hadrosauridae are very well represented in the Judith River deposits. In addition to non-crested forms, such as Edmontosaurus, already considered, are the crested forms represented by three genera, Parasaurolophus, Lambeosaurus and Corythrosaurus. Parasaurolophus was probably the most spectacular of these forms.

Parasaurolophus, based on xxxx (xxxx).
 |
Parasaurolophus has a skull with a truly remarkable nasal crest. As in Saurolophus the nasal passage passes through this crest. In other respects the skeleton is quite similar to Saurolophus. |
Lambeosaurus
Lambeosaurus has a body similar again to that of Saurolophus, but with a very different style of crest.

Lambeosaurus, based on xxxx (xxxx).
 |
The Lambeosaurus skull has a complicated curved nasal crest with a differently shaped nasal passage inside. The genus is named after Charles Lambe. |
Corythrosaurus
Corythrosaurus has again a very different style of crest, in this case similar in shape to a Corinthian helmet - hence the name.

Corythrosaurus, based on xxxx (xxxx).
 |
The Corythrosaurus skull has a complicated curved nasal crest with a differently shaped nasal passage inside. |
Corythrosaurus is represneted by several specimens in nearly perfect articulation, with some skin impression preservation. Given the fine state of preservation is does not take much all that imagination to reconstruct them in life position.

Corythrosaurus skeleton as it appears still imbedded in its rock matrix, based on xxxx (xxxx).
 |
In juveniles the crest is small in size and apparently as individuals grew the crest grew disproportionally larger. |
| In addition, two slightly different crest types seem to present within each type. Dodson argued that these are different sexes. But what function could the crests serve? |  |
| It has been suggested that the crests were weapons, but
they are made of paper-thin bone. It was also suggested that the crests
might serve as some kind of snorkel. There is no opening at the end, however.
Finally, the crests have been interpreted as some kind of extra air supply,
which seems very unlikely, because of its minute volume
In contrast with these seeming quite silly speculations, Weishample (1991) has argued that both the growth pattern and the sexual dimorphism implies a role in species identification and sexual selection. Weishample (1991) hypothesized that the crests, which always carried the nasal passage, were resonating organs that in some way enhanced vocalizations and served in mate recognition and interspecific display. To test his hypothesis, Weishample constructed models of the crests and produced noises with them that were indeed enhanced in different ways depending on their shape. Thus, the hypothesis was corroborated, and its serves as the best explanation of the crest variation to date. |
|
Stegoceras and Pachycephalosaurus
Marginocephalians are extraordinarily well represented in the Judith River beds. Two genera of pachycephalosaurs are present. Stegoceras is a small form similar to Prenocephale from Mongolia, while Pachycephalosaurus is considerably larger and has a very spiky cranium.

Stegoceras skeleton mounted in the Academy of Natural Sciences of Philadelphia.
| In contrast to Stegoceras, the much larger Pachycephalosaurus, has a skull (see right) that has a series of well developed spikes around the margin. | 
Skull of Pachycephalosaurus on display at the North Carolina Museum of Natural Science |
| The Neoceratopsians Chasmosaurus, Centrosaurus,Monoclonius,
and Styracosaurus
Ceratopsians, specifically neoceratopsian are extraordinarily diverse in the Judith River assemblages. Here we will look at just a few that differe primarily in the form of their crests and horns, but otherwse have post-cranial skeltons similar to Triceratops |

Cranium of Chasmosaurus at the Tyrell Museum. |
 |
Chasmosaurus, Centrosaurus, Monoclonius, and Styracosaurus
have very different styles of horns and frills. This seemingly bizarre
assemblage of skull types actually recalls the array of horn styles seen
in antelopes living in East Africa. Again mate selection and dominance
displays and combat probably played a critical role.
On the other hand, it seems almost certain that the horns were also used for defense against predators such as tyrannosaurs. In fact, various neoceratopsian skeletal elements have been found with tooth marks that match those of tyrannosaurs, so they did have something to defend against. |
A diverse assemblage of Coelurosauria are present in the Judith River including ornithomimids, troodontids, tyrannosaurs, and oviraptorids. First we will consider the Arctometatarsalia, specifically the Ornithomimosauria, of the Bullatosauria.
Two ornithomimids are Struthiomimus altus and Dromiceomimus

Skeleton of Struthiomimus altus as displayed in the old hall of Cretaceous Dinosaurs in the American Museum of Natural History/

Dromiceomimus skull as it was displayed at the American Museum of Natural History. |
Dromiceomimus is rather similar to Struthiomimus. Its skull (left) is shows the typical ornithomimid condition of being edentulous and having very large eye sockets with a large brain case (bullatosaurian features). |
Troodontids, namely Troodon formosus and Sauroidithoidesare
also present.
Albertosaurus and Daspletosaurus
The Tyrannosauria are abundant in the Judith River beds. Albertosaurus is the best known form and is represented by excellent skeletons. In most ways it is very similar to Tyrannosaurus, but is smaller and more delicate - if you can say that of a tyrannosaur.

Skeleton of Albertosaurus mounted in the Tyrrell Museum, Alberta.
 |
A particularly realistic reconstruction of Albertosaurus is on display at the Tyrrell museum. It probably had a least a few feathers here or there in addition to the scales shown here. |
| Another tyrannosaurid is Daspletosaurus with a distinctively different looking skull (right). |  |
An oviraptorid Caenagnathusis presnt, with a skull not quite
as bizzar as Oviraptor itself.
Dromeosaurus
The deinonychosaurian maniraptoran Dromeosaurus is present as good skeletons.

Skeleton of Dromeosaurus mounted at the Tyrell Museum.
Dromeosaurus is similar to both Velociraptor and Deinonychus,
but has a stouter, more boxy head.
While most of the dinosaur material from the marine beds consist of scraps, there is a surprising diversity of forms. In addition, an excellent Cretaceous-Tertiary boundary section is presently exposed.

Reconstructtion of Gyposaurus.
| Hadrosaurus foulkihas already been described, but at least one other hadrosaur is present. Originally described as Hadrosaurus minor by Colbert (1948) this form actually seems to be a member of the genus Gyposaurus. It was found in beds immediately below the Cretaceous-Tertiary boundary in the last remaining active marl pit in Eastern North America. |
Another ornithischian is represented by a scrap of ankylosaur armor.
Fragmentary remains of ornithomimids (Coelosaurus antiquus of
Cope) and perhaps tyrannosaurs (Dryptosaurus aqilunguis of Cope
and Albertosaurus) are present (see reconstruction above).
Studies by Russell (1984) and Sheehan and Fastovsky (1992) detect no
statistically significant reduction in total dinosaurian diversity through
the late Cretaceous. The matter is still highly controversial, however.
Bakker, R. T., 1986, The Dinosaur Heresies. Kensington Publishing, New York, 481 p.
Colbert, E. H. 1948 A hadrosaurian dinosaur from New Jersey. Proceeding of the Aademy of Natural Sciences of Philadephia v. 100, p. 23-37.
Dodson, P., 1975, Taxonomic implications of relative growth in lambeosaurine hadrosaurs. Systematic Zoology v. 24, p. 37-54.
Dodson, P., 1996, The Horned Dinosaurs. Princeton University Press, Princeton, 346 p.
Fisher, P. E., Russell, D. A., Stoskopf,M. K., Barrick, R. E., Hammer, M., and Kuzmitz, A. A., 2000, Cardiovascular evidence for an intermediate or higher metabolic rate in and ornithiscian dinosaur. Science 288, p. 503-505.
Horner, J. R. and Makela, R. 1979. Nest of juveniles provides evidence of family structure among dinosaurs. Nature 282: 296-298
Horner, J. R. 1982. Evidence of colonial nesting and 'site fidelity' among ornithischian dinosaurs. Nature 297: 675-676.
Horner, J. R. 1983. Cranial osteology and morphology of the type specimen of Maiasaura peeblesorum (Ornithischia: Hadrosauridae), with discussion of its phylogenetic position. Journal Vertebrate Paleontology 3(1): 29-38.
Horner, J. R. 1987. Ecologic and behavioral implications derived from a dinosaur nesting site. In Dinosaurs Past and Present, Vol. II, S. J. Czerkas and E. C. Olson (Eds.), Univ. Washington Press (Seattle). 51-63.
Horner, J. R. and Lessam, D., 1993, The Complete T. rex. Simon & Schuster, New York, 239 p.
Lockley, M. and Hunt, A. P., 1995, Dinosaur Tracks and Other Fossil Footprints of the Western United States. Columbia University Press, 338 p.
Paul, G. S., 1988, Predatory Dinosaur of ther World. New York Academy of Sciences, New York, 464 p.
Preston, D. J., 1986, Dinosaurs in the Attic. Ballantine Books, New York, 308 p.
Russell, D. A., 1984, The gradual decline of the dinosaurs; fact or fallacy?. Nature v. 307, p. 360-361.
Sheehan, P. M. and Fastovsky, D. E., 1992, Major extinctions of land-dwelling vertebrates at the Cretaceous-Tertiary boundary, eastern Montana. Geology v. 20, p. 556-560.
Varricchio, D. J. and Horner, J. R. 1993. Hadrosaurid and Lambeosaurid bone beds from the Upper Cretaceous Two Medicine Formation of Montana: taphonomic and biologic implications. Canadian Journal of Earth Sciences, 30: 997-1006.
Varricchio, D. J., F. Jackson, J. Borlowski, and J. R. Horner. 1997. Nest and egg clutches of the dinosaur Troodon formosus and the evolution of avian reproductive traits. Nature 385:247-249.
Webb, Grahame J.W. and Carl Gans. 1982. Galloping in Crocodylus johnstoni - A reflection of terrestrial activity?. Records of the Australian Museum v. 34, p. 607-618.
Weishample, D. B., 1981, Acoustic analysis of potential vocalization in lameosaurine dinosaurs (Reptilia: Ornithischia). Paleobiology, v. 7, p. 252-261.
Weishample, D. B., 1984, Evolution of jaw mechanics in ornithopod dinosaurs. Advances in Anatomy, Cell Biology, and Cell Biology v. 87, p. 1-110.
Weishampel, D. B. and Horner, J. R. 1990. Hadrosauridae. Chap. 26 in
The Dinosauria, D. B. Weishampel, P. Dodson and H. Osmolska (Eds.), University
of California Press (Berkeley). 534-561.
Go to the master cladograms: http://rainbow.ldeo.columbia.edu/courses/v1001/masterclads.html