Back to Lecture
22
BACK TO DINOSAURS 2001 HOME PAGE
UNDER CONSTRUCTION
Lecture 23 - Hot Blooded Dinosaurs I - Tracking Dinosaurs
This begins two lectures on dinosaurian metabolism, the first of which
looks at aspects of dinosaur metabolsim and behavior that can be extracted
from the tracks of reptiles.
First however we should consider what aspects of metabolism we are interested
in and how we want to define our questions.
Bob Bakker framed the most provocative question on dinsaurian
metabolism by asking, "were dinosaurs hot-blooded?". By this he meant,
did dinosaurs have a metabolism closer to that of mammals and birds or
did they have one much more like living reptiles?
We need to be more specific, however, and need to focus in on what we
mean by "hot-blooded". Really Bakker meant "warm blooded", but what does
that mean? Many lizards have blood as warm as ours, but are still regarded
as "cold blooded". We can clarify this by breaking temperature regulation
into two catagories:
1) the source of heat to keep warm:
animals can be ectothermic - which means they obtain
heat from their environment,
or they can be endothermic - which means they can produce
their own heat.
2) their temperature variability:
animals can be poikilothermic - having a body temperature
that fluctuates over a fairly wide range,
or they can be homeothermic - having body temperatures
that don't fluctuate much at all.
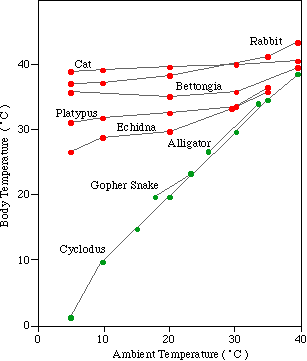 |
We are homeothermic endotherms, that is our body heat is produced by
our own metabolism, as are other mammals (to a greater or lesser extent)
and birds. Lizards are poikilothermic ectotherms, so their temperature
fluctates greatly and they warm up by seeking a warm place, as are crocodilians.
The differing response of the body temperatures of different kinds
of animals can be seen on the graph on the left. Mammals (red) change their
body temperature little with increasing ambient (surrounding air) temperature.
The bodies of reptiles (green), on the other hand, change in proportion
to the ambient temperature.
But Bakker was looking at much more than just temperature regulation
per
se. He was also looking at what we see associated with bird and mammal
style thermoregulation - high activity levels and agility. |
So first we will look at activity levels and then relate this back to
temperature regulation and the general question of dinosaurian metabolism.
Tracking Dinosaurs and The Study of Footprints - Ichnology
There is no branch of detective science so important and
so much neglected as the art of tracing footprints.
A. Conan Doyle 1861
(Study in Scarlet)
One area of dinosaur paleontology that can provide evidence on activity
levels are footprints. Tracks are the only record we have of dinosaurs
in motion and thus they can provide clues about behavior and inferentially,
metabolism.
Tracks can be preserved in a remarkable variety of ways. One common
way is for the tracks to be made in regular sticky mud. These tracks are
often poorly preserved because of partial collapse of the mud back into
the track and mud adhering to the foot.
| The sorts of tracks made in sticky, new mud rarely show much anatomy,
and they are typical of most really large tracks as well as quite a few
small ones. On the right is a modern lizard trackway made in mud that had
been deposited the night before by a flash flood along a river. Note the
lack of detail. This is the style of track most of us make when walking
in mud.

|

Lizard tracks, Red River, Palo Duro Canyon, TX |
Trackway of Rhynchosauroides hyperbates. |
However, some fossil tracks are exquisitely preserved with almost unbelievable
detail. Above left is an impression of a manus of the lizard-like form
Rhynchosauroides
hyperbates from Late Triassic age strata of Pennsylvania (Newark basin).
This track comes from a very long series (left) in which the animal
actually sat down several times (below).

Impression of chest of Rhynchosauroides hyperbates.
The horizontal rows of scales are from the belly and the two patches
of scales above that area was made by the animal's pectoral muscles. This
pattern is nearly identical to what we see on many modern lizards. However,
we don't not know the kind of animal that made these Triassic tracks. It
could have been a lizard, but we don't know.
Obviously, we can learn a lot about the trackmaker's anatomy from
these kinds of tracks. That is not necessarily true of the tracks from
Palo Duro Canyon we looked at above. |
Why is it that these tracks are preserved so well, but the ones made
in fresh mud are so bad? A clue can be gleamed from looking not at newly
deposited mud, but rather at "old" mud.
| On the right are tracks a large number of birds and I made on the shores
of a little pond in Texas. Although my tracks are deep (below) the impression
is very clear, as are those of the birds.
This is because of a coating of algae and bacteria that not only
binds the surface together, but also act as a parting medium, preventing
our feet from sticking to the mud.

You can seen the greenish coating on the damp mud. This algal and bacterial
coating also slows the drying of the mud. |
 |
Any given real track can be thought of as falling somewhere in between
three "end members" of track types. These end members are - 1) tracks whose
form faithfully reflect the trackmaker's anatomy; 2) tracks whose form
is a result of the motion of the foot (or body) through or on the mud,
which we call kinematics; and 3) tracks whose form is a result of the nature
of the substrate.
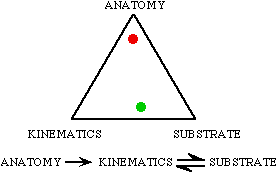 |
We can imagine these three end-members forming the corners of a triangle
- a kind of "space" into which a given track somewhere falls. This kind
of diagram is called a ternary diagram and it is a type often used by mineralogists
and geochemists.
Here the red dot represents the position of the Rhynchosauroides
hyperbates tracks, while the green dot represents the modern lizard
tracks from Palo Duro Canyon, TX. The Rhynchosauroides hyperbates
tracks reflect mostly the anatomy of the trackmaker, while the modern lizard
tracks reflect the interaction between the animal's motions (kinematics)
and the substrate.
The latter point illustrates an asymmetry between the levels of
interactions of the making of tracks. Obviously, the nature of the substrate
(mud) influences the animal's kinematics and the kinematics influence how
the substrate yields; however, neither substrate nor the animal's movements
can change the actual anatomy of the animal - except through natural selection. |
So providing we can get a track that shows anatomy, how do we compare
it to fossil animals known only from their skeletons, not the soft tissue
of their feet?
| The problem is well illustrated from the trackways of the non-dinosaurian
track, Chirotherium, which we saw earlier (and to the right).
Chirotherium means, literally, hand animal and refers to the
way the impression of digit V of the hand sticks out like a thumb. This
bit of anatomy in the track posed a difficulty for the reconstruction of
the skeleton, because no skeleton has a digit V of the pes that sticks
out in such a manner. In fact, no skeleton of any modern reptile does that
either.
However, Kevin Padian (University of California at Berkeley) and
I had a suspicion that it was commonplace for sauropsid feet to interact
with the ground in this way. In 1983 we set out to test this hypothesis
by means of an experiment.
With the cooperation of the San
Diego Zoo, we enlisted the use of a large 13 year old male Komodo
Dragon or Ora (Varanus komodoensis), native to the island of
Komodo in Indonesia.
He was, at that time, 2.54 m (8 ft, 4 in) long and weighed 82.5 kg (~182
lb). We also looked at a smaller female (below) that was 1.8 m and 26.25
kg as well, because she was blind and could be handled! Komodo Dragons
are monitor lizards and are the largest fully terrestrial reptiles alive
today. However our zoo male was about twice as meaty as wild ones.

The female Komodo Dragon. |
 |
The foot has very primitive sauropsid proportions with digit IV being
the longest. Digit V points forward, as it does in modern skeletons and
in fossil lizard and even archosauromorph skeletons.
|

One of the keepers enticed the male to walk on the clay bed with a tasty
dead rat on the end of a stick (right). Notice the large amount of saliva
dripping from the ora's mouth. This saliva is loaded with bacteria that
are instrumental in producing massive infections that the wild Komodo Dragons
use to bring down larger prey. |
In order to test our hypothesis we had to convince the male ora to
make some footprints for us. We made a bed of terra cotta clay and smeared
glycerin on it to act as a separator (left). The zoo keepers arranged barriers
to keep the ora on the clay bed.

|
| Unfortunately, on this go-round the ora pretty much ignored the ladder
barricade and walked on it producing a half-trackway (right). So we had
to level the clay. We really did not need the glycerin, because the ora's
saliva was all over the clay and made an excellent separator. Fortunately,
we had no cuts on our hands!

|

The Komodo Dragon was kept at bay for a while and then turned
around, again using the rat as bait (left). |
| This time (above) he made a perfect trackway and was allowed to keep
his tidbit.

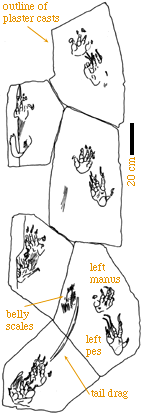
On the right is a plaster cast of a manus and pes set of tracks. The
manus track is in front of the pes. Because this is a plaster cast of a
left set of tracks, the tracks are in mirror image and digit V is on the
right.
Note the scale impressions and that the claws (and ther drag marks)
are longer than they would be for a wild animal.
These tracks reflect the anatomy of the trackmaker very well.
Digit V, however, sticks out laterally, much like in Chirotherium.
Thus the thumb-like shape of digit V in Chirotherium is a primitive
feature (at the level of the Diapsida) and reflects nothing particularly
unusual in foot structure. |
 |
We then produced an outline drawing of the manus-pes set (below middle)
and then reconstructed the skeleton of the hand and foot based on the assumption
that the pads underlie the articulations between bones. The result (bottom
right) compares well to the skeleton of an ora. This is the process we
can go through to compare well-preserved small-to-medium sized dinosaur
tracks to the skeletons of potential trackmakers.
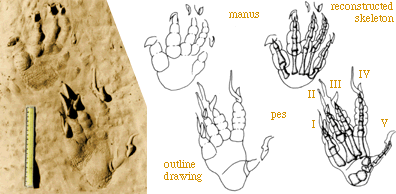
 |
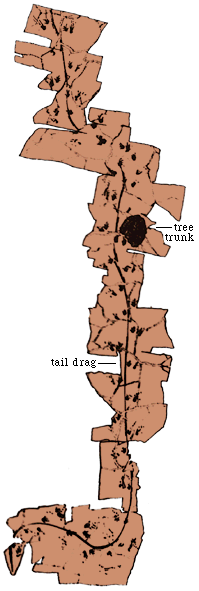
We can also learn something about the gait and even perhaps breathing
mode by looking at the trackway (left).
Note that the trackway is very broad, not at all like the trackway
of Chirotherium. In addition, there is a tail drag that seems to
indicate a sinuous motion to the body and certainly a sprawling posture
and gait. This sinuous motion can also be seen in the photograph of the
walking ora (below)

Such motion in lizards and other more primitive tetrapods alternately
compresses each lung, with the result that lizards cannot breath while
walking and running. They must stop to breath or while walking breath between
steps. This is not true of birds, and presumably dinosaurs. It is also
not true of mammals. Lizards exhibits the so called Carrier's
constraint, the inherited fish-like motion of their ancestors that
limits breathing while walking or (for tetrapods) swimming.
The maximum sprinting range of Komodo Dragons is thus about 10
m (Cowen,
1997).
Crocodiles in a gallop compress their lungs symmetrically and
thus presumably can breathe. But while swimming they undulate their bodies
side to side and hence are under "Carrier's constraint" and can only swim
fast for very short spurts. The trackways of birds and dinosaurs shows
that dinosaurs did not move sinuously while walking or running and thus
could breathe while running (see below). |
Dinosaur Sociality from Tracks
Crude Notes Follow.
II. Let's look at running - Getting at activity levels.
A. Footprints only really indication of how dinosaurs moved
and it is reasonable
to ask if they can tell us about activity.
1. Posture obvious from trackways - but activity levels not.
B. It is possible to get speed from tracks.
C. When you run your pace and stride increase suggesting a way.
Slide of Running pg. 3.7 MCN Alex.
1. Ship designers need way of making model
The way gravity influences motion is not proportional to length.
2. There is a relation called dynamic similarity which takes
this into account.
3. This value tends to vary in proportion for different size ships.
4. R. McNeill Alexander - has worked out a similar relationship for
running tetrapods
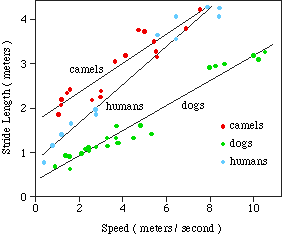
Relationship Between Stride and Speed
5. Suggested relationship.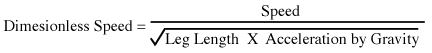
Now we can graph dimensionless speed against relative stride
as follows:
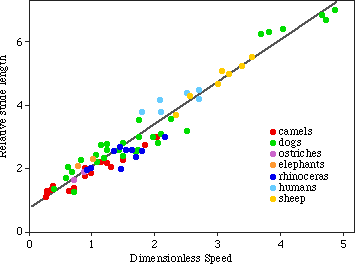
6. We can solve for speed given this relationship.
7. But we still have to get leg length from a footprint.
8. What McNeil Alexannder means by leg length is actual hip height.
Hip height scales to foot length for dinosaurs no matter what the size
or whether they are bipedal or quadrupedal.
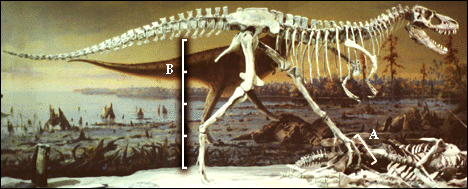 |
| This skelton of Albertosaurus at the Tyrell Museum shows the
aproximate relationship between the phalangeal length (that part of the
foot leaving a footprint - i. e. foot length) and the hip height. |
LEG = 4 X FOOT LENGTH (to metatarsals)
9. Now we have all the information we need to go from a trackway
to an estimate of speed.
From a variety of dinosaur tracks of different ages we get:
| Locality |
Est. Leg Length |
Speed (km/hr) |
Gait |
| large theropod |
2.0 |
8 |
walk |
| small theropod |
1.0 |
13 |
run |
| large sauropod |
3.0 |
3.6 |
walk |
| small sauropod |
1.5 |
4.0 |
walk |
| ornithopods |
0.14-1.6 |
15.5-17.3 |
run |
| small theropods |
0.13-0.22 |
11-12.6 |
run |
| small theropods |
1.0 |
30-40 |
fast run |
| human runner |
1.1 |
36 |
sprint |
| race horse |
1.6 |
61.2 |
fast run |
| ostrich |
1.5 |
54 |
fast run |
Thus:
Most dinosaur tracks are thus of walking animals.
But some could run fast, as fast as the fastest mammals.
So this shows dinosaurs could move as fast as fastest
mammals.
III. Getting at metabolism from dinosaurian structural parallels.
A. Now we can see that dinosaurs could probably run very fast,
but so can some reptiles for short distances.
B. Structurally, however, they are mammal or bird-like.
1. Struthiomimus - Ostrich.
2. Deinonychus - Archeopteryx.
3. Hadrosaur - Elk.
C. Large animals have similar structural requirements.
1. Sauropod and elephant.
IV. Internal Constancy vs. Variability of Temp.
A. What about heat production?
1. Resting metabolic rate greater in mammals and birds than
in reptiles.
SLIDE OF RESTING METABOLIC RATE FROM MC A.
2. Size confers some thermal intertia.
3. Surface area is where heat is lost.
4. Surface area increases as the square of linear dimension.
5. But volume increases as the cube.
6. Therefore for things of similar shape, large things lose heat slower.
7. Big animals have great thermal inertia.
So big dinos could have been inertial
homeotherms.
8. Also this means that large things with high metabolic rates need
to dump heat.
9. But small things like shrews need to conserve it always.
10. Sometimes birds with good insulation need to cool - air sac system.
|
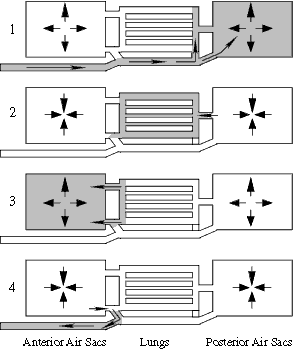
How the air sac system in birds works
Unlike the lungs of non-birds, the lungs of birds are not blind sacs,
but rather tubes through which air moves in a one-way direction.
Step 1: The air sacs inflate as the body cavity expands, a mass
of air (gray) is drawn into the posterior sacs.
Step 2. As the body cavity contracts, the mass of air in the
posterior sacs is pushed into the lungs where gas exchange takes place.
Step 3. As the body cavity expands again, the airs sacs expand
and that mass of air, moves into the anterior sacs. At the same time another
mass of air (not shown) is drawn into the posterior sac.
Step 4. With the next contraction the original mass of air is
exhaled and the new mass (not shown) moves into the lungs,
In this way there is a non-stop flow of air through the lungs.
These air sacs also serve as cooling devices, much like panting is used
by dogs to cool them off.
Adapted from Welty (1975).
|
 |
11. Air sac system present in giant sauropods - probably for cooling.
B. Of course, if you have high metabolic rates you need to eat more food.
SLIDE OF BAKKERS DOG VS LIZARD PG 376
1. Thus we would expect the same number of prey animals to
be able to feed a larger number of low metabolic rate animals than high
ones.
2. In fact if we look at the distribution of predator/prey ratios we
can see that dinosaurs have a lower predator/prey ratio than eocene mammals.
SLIDE OF PREDATOR/PREY RATIOS DRAWING FROM BAK PG 351
3. Thus it evidently took many, many herbivorous dinosaurs
to feed one carnivorous one.
V. Activity Level vs. Bone Strength.
A. It has long been observed that the structure of the bones
in mammals and birds is different than that seen in living reptiles.
ADAPTED SLIDE FROM BAKKER PG 352
B. Haversian system present in birds and mammals.
SLIDE OF HAVERSIAN SYSTEM
EXPLAIN CROMPTON'S ARGUMENT
VI. Phylogenetics.
Shared derived characters of dinosaurs and birds.
Four-chambered vs. three-chambered heart.
VII. Summary
A. Dinosaurs could run very fast.
B. Dinosaurs were built for long periods of activity.
C. Large dinosaurs could have easily been inertial homeotherms
but small ones had to produce extra heat or would have to adapt their behavior
to receive it from environment.
D. Large dinosaurs needed to lose heat.
E. Predator/prey ratios suggest that carnivorous dinosaurs
needed as much food as carnivorous mammals.
F. Dinosaur bones remodelled as often as mammal and bird
bones suggesting high activity levels.
G. Birds are dinosaurs and may have inherited their metabolic
systems.
VIII. Conclusion.
At least some dinosaurs were as active and "warm blooded"
as any mammal or bird.
And so although we can't go back in time and measure
them we can infer, make predictions, and test them in the fossil record.
Note that combined with their locomotor adaptations it
becomes easy to see why dinosaurs remained dominant for so long over mammals.
References cited
Welty, J. C. 1975. The Life of Birds, W. B. Saunders & Co,
623 p.
Go to Lecture 24
BACK TO SYLLABUS
BACK TO THE DINOSAUR AND THE HISTORY OF LIFE
HOME PAGE