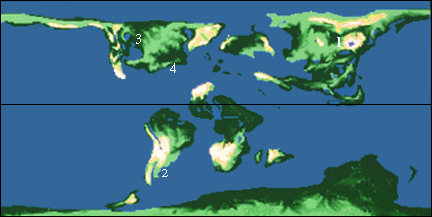
1, Mongolia; 2, Argentina; 3, Western North America, 4; New Jersey.
BACK TO DINOSAURS 2001 HOME PAGE
During the Late Cretaceous the final break up of Pangea continued with the rifting of Eurasia from North America and Greenland, and India, Australia, and Antarctica from Africa. The high rates of formation of new oceanic crust resulted in significant belt of relatively warm (and hence unusually low density) oceanic crust around the spreading ridges. These low density bands of lithosphere floated higher on the asthernosphere than cooler crust and thus displaced ocean waters onto low-lying parts of the continents. The Late Cretaceous is thus a time of very high sea level and vast excursions of marine waters into the continents creating vast epicontinental seas. These fragmented the areas above water into relatively small island continents.
1, Mongolia; 2, Argentina; 3, Western North America, 4; New Jersey.
The rapid rates of oceanic crust formation also undoubtedly pumped unusually large amounts of CO2 into the atmosphere. Perhaps as a consequence, this CO2 ended up being removed from the atmosphere as CaCO3 (Calcium Carbonate) much of which consisted of chalk (see Lecture 18).
Based on recent work, CO2 seems to have been declining during the Late Cretaceous, as may have global average temperatures - although there is little evidence of significant polar ice. The reason for the CO2 decline is controversial, but may be related to the continued evolutionary and ecological radiation of the flowering plants, with concomitant increased continental weathering rates.
| In 1908 Henry Fairfield Osborn, a famous mammalian vertebrate paleontologist became president of the American Museum of Natural History in New York City. He believed he that origin of both humans and dinosaurs lie in Central Asia, largely because he thought that region lay in the center of dispersal of seemingly similar types of humans and dinosaurs. But during the turn of the century most of Asia, especially central Asia, was very poorly known to Western Science, especially paleontology, so Osborn was free to speculate at will. It was known, however, that Mongolia (alternately controlled by Russia and China) has vast expanses of desert, specifically the Gobi desert, a vast region with badlands with excellent rock outcrops that would be the area to look for "missing links", were they to be found in Asia at all. |  |
Roy Chapman Andrews was an ambitious researcher who had worked his way up at the American Museum from floor washer to professional mammalogist. In 1920 he proposed to Osborn that he lead a massive expedition to Outer Mongolia, not only to search for the "missing link" between humans and apes but also to conduct a general scientific study of the region, which was indeed very poorly known.
Although it was potentially very risky, Osborn enthusiastically agreed, and private funds were raised for what became known as the Central Asiatic Expedition, which departed for Beijing in 1922.
According to Preston (1986), Roy Chapman Andrews was the real basis for the fictitious Indian Jones character of "Raiders of the Lost Ark", played by Harrison Ford. The picture on the left from the frontispiece of Andrews' autobiography, is described by Preston (p. 120-121) as follows.
 |
"[The picture] shows [Andrews] sitting on a dessert hillock. He holds a 6.5 mm Mannlicher rifle in his right hand, and wears a cartridge belt loaded with bullets around his waist. On his feet are dusty leather jackboots . His rumpled wool shirt is rolled up at the sleeves. A worn ranger hat , sporting a pheasant feather, is perched on his head, cocked at an angle. The man is pictured in profile, his hard, clean jaw slightly elevated , his pale eyes gazing off into the distance. Behind him rise the Flaming Cliffs of Shabarakh Usu [Bayn Dzak], in the heart of the Gobi Desert." |
The Central Asiatic Expedition of (1921-1930) and following research projects, notably the Polish Mongolian Expedition (xxxx-xxxx), the Canadian-Chinese Dinosaur Project (xxxx), and the recent American-Mongolian expeditions (199x-present) produced one of the greatest assemblages of the dinosaurs from the world.
The most famous Late Cretaceous dinosaur-bearing formations are the Djadochta and Nemegt, both containing abundant eolian (wind) and water-laid deposits. Below the best known of the dinosaurs from these two formations are briefly described along with their phylogenetic relationships.

Based on Carpenter (1997).
Pinacosaurus is a common dinosaur in the Mongolian deposits. It is an ankylosaurid (member of the Family Ankylosauridae) as opposed to a nodosaurian as we saw in Sauropelta from the Cloverly in the previous lecture.
Ankylosaurids differ from nodosaurians in retention of a primitive configuration of the skull roof (e. g. there are still paired plates). However, ankylosaurids also have the novel shared derived character of having a bony club at the end of the tail (above). Ankylosaurids, such as Pinacosaurus also tend to have broader skulls than nodosaurids and tend to have spikes on the rear covered of their box-like skulls.
Saurolophus angustirostis
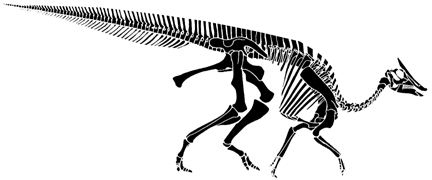
Postcrania based loosely on Parasaurolophus from Brett-Surman (1997) and skull after Norman and Weishample (1990).
Saurolophus is a medium-sized hadrosaurian (member of the Hadrosauria) ornithopod from Mongolia. It is also a crested hadrosaur, which means that it has a crest composed largely of the nasal bones, a condition that seems to have evolved independently among different hadrosaur groups (hence having a crest has little phylogenetic significance.
Hadrosaurian ornithopods are among the most common of all dinosaurs in the Late Cretaceous, in general, although not especially so in Mongolia. In many ways they are similar to iguanodontids, except they tend to have a more superficially duck-like snout - thus the common name "duck-billed" dinosaurs.
| The duck-like snout of Saurolophus can clearly be seen in the
skull on the right. The portion of the snout lacking teeth was covered
by a keratinous beak (as in birds or turtles).
In this case the crest is small, especially compared with other crested hadrosaurs. Species of Saurolophus are know from Western North America, as well as Central Asia.
|
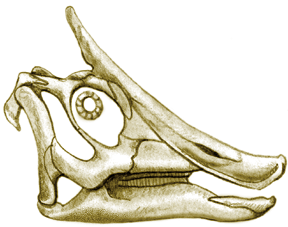
Skull of Saurolophus, after Weishample and Horner (1990)
The shared derived character for the Hadrosauria is the loss of digit I of the manus. This completes a trend seen in the ornithopod of the reduction of digit I starting with the complete digit in hypsilophodontids and Heterodontosaurus, with reduction in Camptosaurus and other iguanontids, culminating in the hadrosaurians. On the left are the manus and pes of a hadrosaur (Lambeosaurus in the Royal Tyrrell Museum in Alberta Canada showing the loss of digit I of the manus. |
Prenocephale prenes
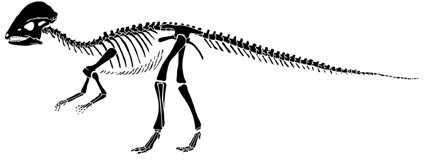
Prenocephale: postcrania based loosely on Stegoceras from Forester and Sereno (1997) with skull of Prenocephale from the same source.
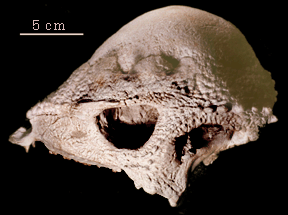
| Prenocephale, is a member of the Pachycephalosauria, the shared
derived character for which is the thickened skull roof, which in this
case makes a massive dome. The Pachycephalosauria, or dome heads, are the
sister group to the Ceratopsia within the Marginocephalia.
In general, pachycephalosaurs tend to be small lightly-built dinosaurs, comparable in size to sheep or antelope (although there are exceptions). Like sheep, it is thought that pachycephalosaurs butted heads during dominance rituals, and the massively thickened skull protected their small brains. The skull (right) of Prenocephale shows the spectacular domed skull roof. Despite its cerebral appearance, the high forehead is all bone. |
 |
Homalocephale calathoceros
| Another pachycephalosaur from Mongolia is Homalocephale. It
is not known from nearly as abundant material as Prenocephale and
it is considerably more primitive.
Unlike Prenocephale, Homalocephale does not have a dome at all. Instead the skull roof is flat and thickened. |
 |
Protoceratops andrewsii
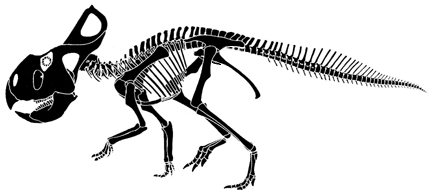
Protoceratops redrawn from Paul (in Dodson, 1997).
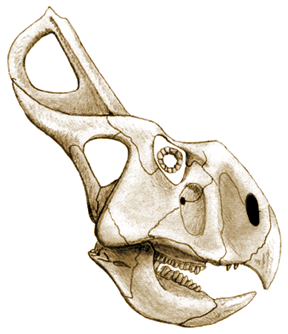 Skull of Protoceratops
andrewsii. Based on Paul (in Dodson, 1997) Skull of Protoceratops
andrewsii. Based on Paul (in Dodson, 1997) |
Protoceratops is the most common dinosaur in the Late Cretaceous
of Mongolia, being represented by hundreds of individuals. Skeletons of
this dinosaur found at Bayn Dzak (the Flaming Cliffs; Djadochta Formation)
contributed greatly to the most spectacular results of the Andrews expeditions.
It was discovered on the first expedition, and on the second expedition
Andrews numerous skulls and skeletons. Enough skeletons were found that
it was possible to look at the growth sequence of embryos to adults and
make a strong case for sexual dimorphism (Dodson, 1976; Dong and Currie,
1993).
Protoceratops is a small neoceratopsian that lacks well developed horns. Its crest is well-developed, but not massive. |
(You can also go to the summary cladograms via this link: http://rainbow.ldeo.columbia.edu/courses/v1001/masterclads.html
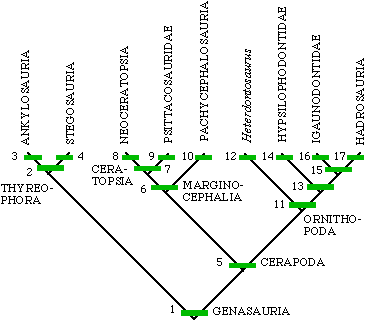
| 1. cheeks
2. armor 3. armor as plates covering back 4. armor as spikes or plates 5. uneven enamel 6. back part of skull makes shelf 7. rostral bone 8. frill 9. reduced hands 10. domed head 11. jaw joint below tooth row 12. elongate hands 13. prepubic process well developed 14. ridge on teeth 15. reduction in manus digits I and V (and large nares) 16. manus digit 1 spike-like |
 |
| While sauropod dinosaurs had certainly declined in prominence since the Late Jurassic, they still were a significant part of the dinosaurian communities in the Late Cretaceous, probably right up to its abrupt end. Nemgtosaurus mongoliensis is a medium sized sauropod from Mongolia, recognizable as a diplodocid by the restriction of teeth to the front of the mouth. | 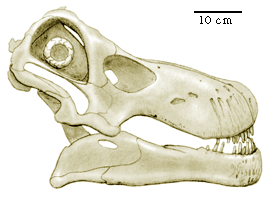 |
Other than the somewhat jutting mandible in Nemgtosaurus mongoliensis the skeleton appears to be of normal diplodocid proportions.
Sauroidithoides junior
The little Sauroidithoides junior is recognizable as a theropod by its three-toed pes, and as a member of the Coelurosauria by its long arms. Sauroidithoides has rather small but numerous teeth, and these teeth sport disproportionally large tubercles along their edges. The orbits are very large and the brain cavity is huge (for a non-bird dinosaur).
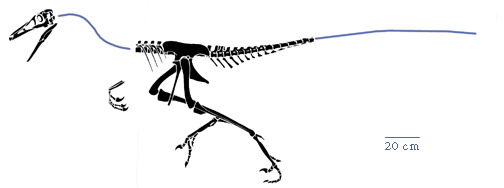 Redrawn from Paul (1988) from his
Troodon mongoliensis based on AMNH 6516.
Redrawn from Paul (1988) from his
Troodon mongoliensis based on AMNH 6516.

Teeth of Sauroidithoides, left, compared with those of Troodon. |
The distinctive teeth of Sauroidithoides are similar to those of Troodon from western North America. This kind of tooth structure is nearly unique among the Dinosauria and is the shared derived character for the Troodontia. These kind of teeth probably served for gripping small prey, such as small lizards, mammals, and insects, rather than for cutting flesh like more typical, and proportionally larger, theropod teeth. |
The reduction in tooth size and well as the proportionally very large brain is also seen in the ostrich mimic dinosaurs, the Ornithomimia (see below). These characters along with others allow the recognition of the group Bullatosauria, consisting of the Troodontia and the Ornithomimia.
| Sauroidithoides also has a distinctive development in the pes also seen in a surprising array of Cretaceous theropods. Specifically, metatarsal III of the pes narrows proximally between metatarsal II and IV to a thin splint. Holtz (xxxx) argues that this feature is a shared derived character uniting the Troodontia with the Ornithomimia and most surprisingly with the Tyrannosauria. This group has been named the Arctometatarsalia, and although we will recognize this group here, it is far from non-controversial. A plausible hypothesis is that this configuration is an adaptation for reducing compressive stresses along the axis of the foot while running. |  |
Gallimimus bullatus

| Gallimimus bullatus is an extremely graceful dinosaur that is characterized by usually having no teeth (edentulous) and by having very long arms in which the manus has three metacarpals of nearly equal length. The latter character is a shared derived character for the Ornithomimia, or "ostrich mimic" dinosaurs, so called because of their rather ostrich-like proportions and skull that must have supported an ostrich-like beak. Note especially the very long tibia and metatarsals relative to the femur, contributing much to its bird-like appearance. | Ornithomimid manus |
Recently an ornithomimid with teeth has been discovered in Spain. Pelicominimus is a seemingly normal ornithomimid except that it has very numerous, very small teeth in both jaws. The condition is reminiscent of the Troodontia and it possible that the latter, if defined only on the basis of its teeth, could be a paraphyletic group. In any case, Pelicominimus shows that the character transformation of the teeth to the edentulous condition seen in most ornithomimids occurred by reduction in size, first, not in number.
Deinocheirus mirificus
 |
The shared derived feature of the Ornithomimia, the three sub-equal metacarpals is seen in a shockingly large, but sadly very incomplete, dinosaur, Deinocheirus mirificus, known only from its arms and shoulder girdle. What the rest of this dinosaur looked like is a mystery, but it seems rather convincingly a member of the ornithomimia. If it were proportionally scaled to Gallimimus, it would be gigantic, larger than Tyrannosaurus rex! |
Tarbosaurus bataar
By far the top carnivore in the Mongolian assemblages was the theropod Tarbosaurus bataar. Tarbosaurus is massively built, with a very large skull on a relatively short neck.

 |
The arms, however, are diminutive, with only two functional fingers. This character is shared with, among others, the famous Tyrannosaurus rex itself, and is the shared derived character for the Tyrannosauria.We will delay the discussion of details of tyrannosaurs until the next lecture (Lecture 20). However, it is important to note that Tarbosaurus does have a typical arctometatarsalian metatarsus (as do all tyrannosaurs) which is the main reason they are now considered coelurosaurs. Tyrannosaurs traditionally were classified as carnosaurs, a practice that was based on shared similarities that are probably adaptations to a large size (i.e. analogous, not homologous features). |  |
Velociraptor mongoliensis
The Maniraptora are very well represented in the Mongolian Late Cretaceous, both members of the Deinonychosauria and Avialia.

Velociraptor mongoliensis, redrawn from Paul (xxxx).
Velociraptor mongoliensis is one of the best known of all of the Mongolian saurischians, and it is famous as well, having starred in both "Jurassic Park" movies. In most ways Velociraptor resembles a more slender Deinonychus and possess, like the latter a well-developed tearing claw on digit II of the pes. The two genera are so similar that Paul (xxxx) attempted to synonomize Deinonychus and Velociraptor, retaining the latter genus name for both forms. This has not been widely followed, however.
 |
The skull is definitely more shallow in Velociraptor than Deinonychus, with a distinctly much more concave dorsal edge seen in all specimens (see left). |
Predation in Velociraptor and deinonychosaurs in general, has been reconstructed as a lunge and with the arms (the rocking motion of the semilunate carpal playing critical role) followed by a push and tear with the hind limbs and the specialized trenchant claw on digit II.
| Amazingly, this hypothesis has been very strongly corroborated by the discovery of a Velociraptor preserved in the act of killing a Protoceratops. This specimen (right), discovered by the Polish-Mongolian expeditions shows a Velociraptor with its hands wrapped onto the head of a Protoceratops while the hind feet of the carnivore are against its belly of the herbivore. Evidently, both were entombed by the collapse of a sand dune. The Velociraptor seemingly could not get away because the Protoceratops had closed its jaws on one of the former's arms. |  |
 |
A critical argument long used against the dinosaurian origin of birds
has been the supposed lack of clavicles in dinosaurs. As we have seen this
is actually not the case. A recently described very nice example of dinosaurian
clavicles is afforded by a specimen described by Norell et al. (xxxx).
Not only are clavicles present, but they are fused into a very bird-like
furcula (left).
Together with the well-developed sternum seen in Velociraptor, the entire shoulder girdle is very similar to that seen in primitive birds. |
Mononychus olecranus
The Avialia are represented by several forms in the Mongolian deposits, but perhaps the most remarkable is Mononychus olecranus, known from relatively complete remains.

According to Norell et al. (199x), who originally described it, Mononychus olecranus is a bird (avialian) because it has a manus that show a fusion of bones seen in birds considerably more advanced than Archaeopteryx. However, the arms of Mononychus are very short and stoutly constructed (below), with only one large pointy digit. Norell et al. (199x) argue that Mononychus was secondarily flightless. That is, it was descended from an ancestor with fully developed flight. The long tail, and structure of the pelvis, suggest that if this hypothesis is true, the bird from which Mononychus was descended was not very advanced.

Oviraptor mongoliensis
Two coelurosaurian groups, the Oviraptoria and the Segnosauria, vie for the most bizarre of Mongolian dinosaur groups. We will consider the Oviraptoria first as represented by its best known representative the eponymousOviraptor mongoliensis.

Oviraptor mongoliensis is a fairly common Mongolian dinosaur. It is named after its supposed habit of stealing eggs of other dinosaur (ova- raptor = egg stealer), an example of which was the little skeleton found on top the eggs by Roy Chapman Andrews and his expedition.
 |
The skull of Oviraptor is truly weird. It is edentulous, and the skull roof is inflated into a large crest full of fenestrae. The palate curves downward, rather than upward as is usual, and in the middle is a pointed tooth-like structure (left). What exactly the peculiar structures on the skull of Oviraptor were for is unknown. |
| The skeleton of Oviraptor is generally maniraptoran-like, although it lacks the typical maniraptoran-type of semilunate carpal and the pubis points forward unlike other bone fide members of the Maniraptora. The tail is incompletely known but seems rather slender. The shoulder girdle has a very well developed furcula and sternum (right). It has long arms and is clearly a coelurosaur. |  |
 |
Norell et al. (1xxx) recently described an additional specimen of Oviraptor found on top of eggs. While the skeleton is fragmentary, the arms and legs are draped around the eggs in exactly the position seen in birds that nest on the ground (right). |
In the nest in which the embryo-bearing Oviraptor eggs were found, Norell et al. (x9xx) also found a partial skull of a baby Velociraptor. Two hypotheses come to mind to explain this association:
In any case, the new discoveries show Oviraptor to have exhibited very bird-like nesting behavior. Norell et al. (1xxx) have reconstructed the position of the nesting Oviraptor just prior to its death as shown below.

The Early Cretaceous Liaoning deposits of China have produced an ovaraptorid, Caudipteryx. It has a fan of feathers on its tail and symmetrical feathers on its arms.

Another ovaraptorid had a shortened tail with distal vertebrae fused into a so-called pygostyle, otherwise seen in advanced birds. Presumably this represents a convergent evolutionary adaptation, because the rest of the skeleton is more primitive than maniraptorans..
It seems fairly obvious, however, that the Oviraptoria represent an aberrant branch of the Coelurosauria with many very bird-like features, both anatomically and behaviorally. How exactly the ovaraptorans fit within the Coelurosauria is not obvious and I place them within an unresolved polytomy with the other groups of coelurosaurs plus Compsognathus (see below).
The Segnosauria
Segnosaurus galbinensis, Erlikosaurus andrewsii, and Therizinosaurus cheloniformes are bizarre saurischians know from fairly incomplete remains (in the case of Therizinosaurus,VERY incomplete), but enough is known to get a good idea of their general structure. They all share unusually large and elongate unguals (terminal phalanges) on the manus. Segnosaurus and Erlikosaurus, at least, have a backwardly projecting pubis suggestive of the Maniraptora, but the neck is long, but massive. The skull has an edentulous, ornithischian-like beak anteriorly, and posteriorly the dentition looks prosauropod like!

Therizinosaurus cheloniformes is based on a single huge claw. Its shape is very similar to the hand claws of segnosaurs. Even if this claw belong to a member of the Segnosauria, which have disproportionally large claws, it would still represent a gigantic dinosaur.
For a time it seemed that the Segnosauria might be descendants of the Prosauropoda, but it seems now that they actually have a fairly extensive suite of coelurosaurian characters. Presently, they appear to be coelurosaurs that have convergently evolved a prosauropod-like habitus. They also appear to have a superficial resemblance in form to a group of ungulate mammals known as the calicotheres, which lived during the Miocene. The structural adaptations seen in the segnosaurs suggests herbivory on trees, although, some have suggested a termite-eating life style, with the huge claws being used anteater-like to rip open their mounds. Like the Ovaraptoria, its relations within the Coelurosauria are uncertain.
(You can also go to the summary cladograms via this link: http://rainbow.ldeo.columbia.edu/courses/v1001/masterclads.html
| 1. three-toed foot
2. digits IV and V lost on hand 3. long arms 4. semilunate carpal 5. fused pelvis 6. large hole in lachrymal bone in skull 7. ?no unique derived characters? 8. giant, hook-like claw on digit II of pes 9. flight feathers 10. ?no unique derived characters? 11. proximal half of metatarsal of digit III pinched 12. teeth lost in front of mouth and very large claws on manus |
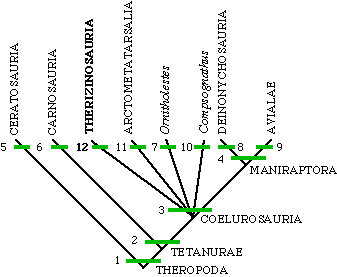 |
Carpenter, K., 1997, Ankylosaurs. In Farlow, J. O. and Brett-Surman, M. K. (eds.) The Complete Dinosaur, Indiana University Press, Bloomington, p. 307-316.
Dodson, P., 1976, Quantitative aspects of relative growth and sexual dimorphism in Protoceratops. Journal of Paleontology v. 50, p. 929-940.
Dodson, P., 1996, The Horned Dinosaurs. Princeton University Press, Princeton, 346 p.
Dong, Z. M. and Currie, P. J., 1993, Protoceratopsian embryos from Inner Mongolia, Peoples Republic of China. Canadian Journal of Earth Science v. 30, 2248-2254.
Forster, C. A. and Sereno, P. C., 1997, Marginocephalians. In Farlow, J. O. and Brett-Surman, M. K. (eds.) The Complete Dinosaur, Indiana University Press, Bloomington, p. 317-329.Norell et al. (199x)
Norell et al. (xxxx)
Norell et al. (1xxx)
Norell et al. (x9xx)
Paul, G., xxxx
Preston, D. J., 1986, Dinosaurs in the Attic. Ballantine Books, New York, 308 p.
Russell (xxxx)
Weishample, D. B. and Horner, J. R., 1990, Hadrosauridae. In Weishample. D. B., Dodson, P., and Osmolska, H. (eds.) The Dinosauria. University of California Press, Berkeley, p. 534-561.
