I. The Triassic World The end-Permian mass extinction brought an end to the Paleozoic Era and marked the beginning of the Mesozoic Era that would last from 245-65 million years ago. This was the "Age of Dinosaurs".
The first period of the Mesozoic Era is the Triassic Period, which lasted from 245 to 202 million years ago.
The name Triassic comes from Germany where it was originally named the Trias in 1834 by Friedrich August Von Alberti (1795-1878) because it is represented by a three-part division of rock types in Germany. It is still called the Trias by many European geologists. The three distinctive formation are (from the bottom up) the Bunter (brown sandstone and red beds), the Muschelkalk, and the Keuper. These turn out to be local rock units that cannot be applied globally.At the end of the Triassic there was another mass extinction, which we will discuss in another lecture.The Triassic was a Hot House world, with no evidence of ice at the poles.The Triassic was critical for Terrestrial life because all of the living dominant groups of tetrapods evolved by its end. These include -Pangea was fully assembled and remained so through the Triassic.
SALAMANDERS
FROGS
TURTLES
DINOSAURS (now surviving as birds)
MAMMALS
CROCODILES
LIZARDS (incl. sphenodonts & snakes)
ALSO PTEROSAURS
ORIGIN OF ANGIOSPERMS (possibly)
In the oceans, ammonites recover, and the first modern corals appear.
At first the worlds' oceans had a remarkably uniform fish fauna - perhaps even at the species level. This was probably a result of the end-Permian mass extinction. Reptiles were at first very rare in the oceans but they became more and more common through time.Seed plants continue their dominance on land. However, the Glossopteris-dominated flora of the southern continents gave way to one dominated by another peculiar seed plant, Dicroidium, and so despite the continuity of the continents, separate floras persisted through the Triassic. In the northern hemisphere, conifers apparently replace Dicroidium and the latter is nowhere to be seen.
 |
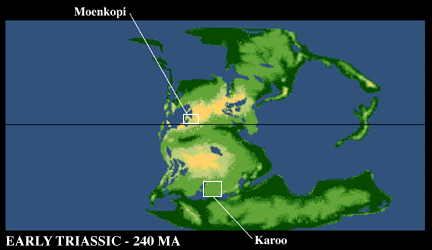
Exposure of red beds of the Moenkopi Formation along Interstate 40 near Winslow, Arizona. Note the red color of most of these rocks, thought to be deposited in an arid to semi-arid coastal plain. This formation is famous for reptile footprints, especially those named Chirotherium |
| Chirotherium is the footprint of an archosauriform, and although at least several species are probably represented they were most likely not very different from Euparkeria. On the right is a slab described by Frank Peabody in 1948 of the natural cast in sandstone of Chirotherium barthi from near Cameron, Arizona (modified from original photo taken by L. F. Brady in Peabody, 1948). This is a species known from the Bunter of Germany and similar-aged strata elsewhere. Scale bar is 20 cm. The name means literally hand animal, and when tracks of this type were originally found in the Early Triassic of England, it was thought that the digit that looks like a thumb was actually the big toe (digit I). In fact it is the little toe (digit V). Charles Lyell thus drew a reconstruction of the Chirotherium trackmaker so that the hind legs crossed as it walked.
Lyell's Reconstruction of the trackmaker of Chirotherium |
 |
However, once the remains of archosauriform "reptiles" began to come to light, it became possible for paleontologists such as Wolfgang Soergel to produce reconstructions (below) that are essentially the same as we would make now.

Reconstruction of Chirotherium by Soergel (1925)
Unfortunately, the Moenkopi is rather poor in actual bone remains, having produced mostly remains of various "amphibians". Other places in the world such as the Ural Mountains in Russia and especially the Karroo basin in southern Africa have produced most of what we know about Early Triassic terrestrial vertebrates.
The unit most famous for bones is called the Beaufort. Beaufort beds have produced a remarkable array of synapsids as well as some important archosauriform "reptiles", notably Euparkeria.
The synapsids are represented by therapsids, distinguished by having a reduced phalangeal formula. The therapsids themselves are divided into two groups, the herbivorous dicynodonts, and the carnivorous cynodonts.Glossopteris was still the dominant southern hemisphere tree during the Early Triassic.
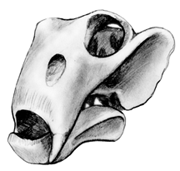
Most abundant was the odd smallish dicynodont, Lystrosaurus. Its overall body shape was rather hippo-like, although it was considerably smaller.
 |
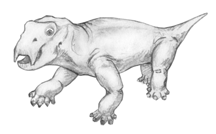 |
| Skull of Lystrosaurus (about 1/3 size) | Reconstruction of Lystrosaurus |
In Lystrosaurus we see the characteristic barrel-shaped body and the reduction of teeth (to two tusks) typical of dicynodonts. There almost certainly was a turtle-like beak as well. In addition to Lystrosaurus, there were a reasonably large number of other smaller and larger herbivores.
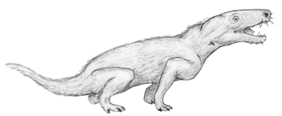
A representative Beaufort cynodont is Thrinaxodon. It has a surprisingly mammal-like skull with a clear reduction in the size of the bones in the back end of the lower jaw.
 |
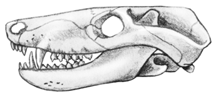 |
| Reconstruction of Thrinaxodon | Skull of Thrinaxodon (about 1/2 size) |
I have reconstructed Thrinaxodon with hair, for which there really is no unequivocal evidence. However in overall Gestalt and in many details there really is no doubt that cynodonts, such as Thrinaxodon, look very mammal-like. There were other cynodonts around during late Beaufort time, some fairly large.
Of course archosauriform "reptiles" were becoming quite common, especially as carnivores. Euparkeria from the Beaufort is an excellent example.
Very early lizards or lizard relatives are also known from Madagascar .
In the oceans, a variety of "reptiles" began to establish themselves and developed clear aquatic adaptations. The most notable were the ichthyosaurs, which developed a very porpoise-like body. Ichthyosaurs may be diapsids, which have lost their lower temporal openings. The earliest known ichthyosaur is Utatsusaurus from Japan.
Below are depicted some of the aquatic reptiles found in finely laminated and organic-rich rocks that formed in these lagoons. Since the Triassic, that part of northern Africa drifted north with the rest of the plate and formed the Alps, in which the rocks now outcrop in the Lake Lugano region of Italy and Switzerland.
|
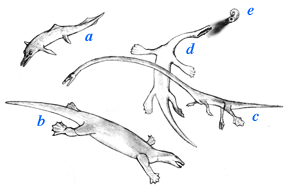
Marine reptiles from Monte San Giorgio:
a, the ichthyosaur Mixosaurus Placodus in this reconstruction was about 2.5 m long |
 |
Early on in the Triassic one or two branches of diapsids lost the lower bar of bone below the lower temporal opening. These groups diversified into a large range of forms, some of which evolved into lizards, others evolved into many forms some of which had aquatic adaptations. The exact relationships of these forms within the Diapsida is controversial, but they almost certainly branch off somewhere between the lizards and the Archosauriformes.
Some of the most bizarre forms are the tanystropheids, particularly Tanystropheus from Europe and Tanytrachelos from the eastern US. Tanystropheus is from marine deposits and has an absurdly long neck, with otherwise rather lizard-like proportions. Tanytrachelos is very much smaller and even more lizard-like, and comes from lake deposits in the eastern US (Newark Supergroup). Tanystropheus and Tanytrachelos have a pair of bones at the base of their tail that evidently served as supports for hemipenes (paired penises present in lizards), only of course in the presumed males.Two closely related groups were the nothosaurs and placodonts. The nothosaurs were mostly small and tended to develop long necks and serpentine proportions. They are the sister group to the plesiosaurs, which become very common in Jurassic and Cretaceous seas. Nothosaurs were probably rather like seals in their habits.
Placodonts had much more chunky proportions, with some developing very turtle-like proportions. One of the more generalized forms was Placodus which probably ate mollusks. Placodonts were probably similar to walruses and marine turtles in their habits. Nothosaurs and placodonts became extinct at the end of the Triassic.
Ichthyosaurs appear in Early Triassic marine deposits. Middle and Late Triassic forms such as Cymbospondylus (10 m long, M. Triassic, Nevada) and Shonisaurus (15 m long, L. Triassic, Nevada) grew enormous. They had very porpoise-like proportions and presumably ecological requirements.
Lizards evolved in the Triassic and became quite common by its end. Some early forms evolved into gliding forms such as Kuehnosaurus from England and Icarosaurus from New Jersey. Both had a gliding membrane supported by vastly expanded ribs. This sort of adaptation to gliding has evolved at least two other times independently: once in the Permian diapsid Coleurosauravus and again in the living lizard Draco. The Triassic forms died out at the end of the Period.
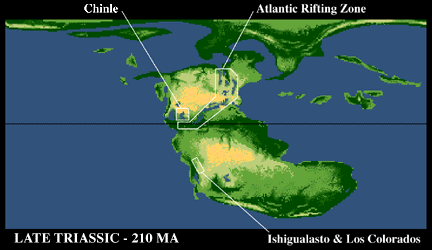
The continents remained united in the supercontinent of Pangea during the Late Triassic. However, the North American and Eurasian Plate (together called Laurasia) began to slowly pull away from the African, South American, and other major plates (collectively called Gondwanaland). In the process a large series of rift valleys formed along the stretching crust that in the Jurassic would become the Central Atlantic Ocean. On the opposite sides of the plates there was convergence. All this slow movement of plates created a much larger array of basins world-wide that preserved Late Triassic (and Early Jurassic), largely continental strata. Therefore we know much more about the Late Triassic than either the Early or Middle Triassic.
As in the Early and Middle Triassic, despite the continuity of the continents, separate Laurasian and Gondwanaland floras persisted through the Late Triassic. Laurasia, remained dominated by conifers, other seed plants and ferns, while Gondwanaland remained dominated by Dicroidium.
Sedimentary rocks of the Ischigualasto Formation are largely drab gray in color and seem to have been deposited in a humid environment. Situated to the east of a subduction zone, the area received ash from volcanoes. This ash has provided a date of 228 million years ago for the Ischigualasto.
| Terrestrial vertebrate fossils are abundant in the Ischigualasto. Most common are the skeletons of advanced synapsids and odd, beaked, archosauriform "reptiles" called rhynchosaurs. Also present are large carnivorous archosauriforms called Rauisuchians and smaller rather different archosauriforms that we will discuss shortly. | 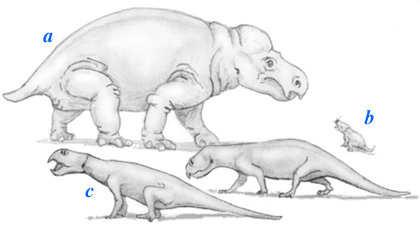 |
However, the stars of the Ischigualasto Formation are clearly the dinosaurs, of which several are represented.
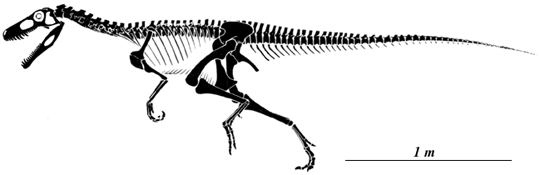
Herrerasaurus ischigualastensis, based on Paul (1988) and Sereno (1993).
Originally described by Reig (1963), Herrerasaurus ischigualastensis has recently been redescribed on the basis of new and very complete material discovered by a team lead by Paul Sereno of the University of Chicago.
The skull of Herrerasaurus is not all that different from other Archosauriformes, such as Euparkeria.
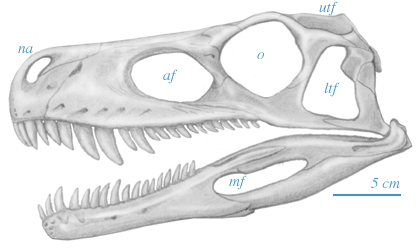 |
Skull of Herrerasaurus ischigualastensis, based on Sereno and Novas (1993). af, antorbital fenestra; ltf, lower temporal fenestra; mf, mandibular fenestra; na, nares; o, orbit; utf, upper temporal fenestra. |
| While the skull has many characters primitive to Archosauriformes in general, the pelvis shows a real dramatic break with the past. Unlike the pelvis seen in other sauropsids, that of Herrerasaurus lacks bone in the center of the acetabulum (hip socket). It turns out that this is the key shared derived character for dinosaurs, and it lets us recognize Owen's group, the Dinosauria. On the right, the Herrerasaurus pelvis is shown in lateral view with anterior being to the right. Abbreviations are; ac, acetabulum; il, ilium; is, ischium; and, p, pubis. Drawing is based on Novas (1993). |
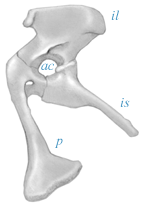 |
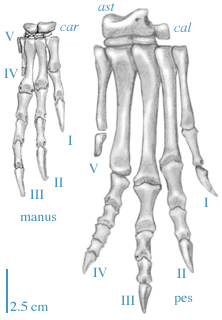 |
The manus (hand) of Herrerasaurus is greatly modified from the primitive Archosauriform condition in the great elongation of digits I, II, and III and the corresponding reduction in digits IV and V. However, this is probably not a dinosaur shared derived character per se, rather it is a shared derived character for a subdivision of the Dinosauria, the Saurischia.
This structure of the manus of Herrerasaurus is extremely important, because it demonstrates the homology of the manual digits within the Theropoda and hence the birds. This point will be discussed in more detail in later lectures.The pes (foot) of Herrerasaurus is not very modified from that seen in the primitive condition for the Archosauriformes, and little different from Euparkeria, except in the ankle. In the ankle the joint with the leg is made between the astragalus plus calcaneum which are attached to the tibia and fibula, respectively, and the distal tarsals, and it is a simple hinge joint. This is a derived character, again not for the Dinosauria, but at a slightly higher level, the Ornithodira, which we will examine in the next lecture. Abbreviations are: ast, astragalus; cal, calcaneum; car, carpals; I-V, digits I through V. Manus modified from Sereno (1993), and pes modified from Novas (1993). |
Novas, F. E., 1993, New information on the systematics and postcranial skeleton of Herrerasaurus ischigualastensis (Theropods: Herrerasauridae) from the Ischigualasto Formation (Upper Triassic) of Argentina. Journal of Vertebrate Paleontology, v. 13, no. 4, p. 400-423.
Paul, G. S., 1988, Predatory Dinosaurs. New York, New York Academy of Sciences, 464 p.
Peabody, F., E., 1948, Reptile and amphibian trackways from the Lower Triassic Moenkopi Formation of Arizona and Utah. University of California Bulletin of Geological Science, v. 28, p. 295-468.
Sereno, P. C., 1993, The pectoral girdle and forelimb of the basal theropod Herrerasaurus ischigualastensis. Journal of Vertebrate Paleontology, v. 13, no. 4, p. 425-450.
Sereno, P. C. and Novas, F. E., 1993, The skull and neck of the basal theropod Herrerasaurus ischigualastensis. Journal of Vertebrate Paleontology, v. 13, no. 4, p. 451-476.
Soergel, W., 1925, Die Fährten der Chirotheria. Jena, G. Fischer, 92 p.
