BACK TO DINOSAURS 1997 HOME PAGE
Great Triassic Assemblages Pt 1 - The Chinle and Newark
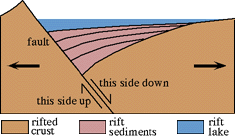
Stretching of the crust produced a broad series of rift basins in the Atlantic Rifting Zone. In Eastern North America, these rifts are preserved from Newfoundland to eastern Texas. The exposed strata of these eastern US rifts are termed the Newark Supergroup, after Newark, New Jersey. The thickness of Triassic and Early Jurassic rocks preserved in these rifts can exceed 7 km.
| Typical Rift Basin | Typical Outcrop of Triassic Rift Strata, Newark basin, Rutherford, NJ |
|---|---|
 |
 |
Newark Supergroup strata are famous for the large variety of fossils found in strata formed mostly in ephemeral to deep lakes. Some examples are shown in this photo album.
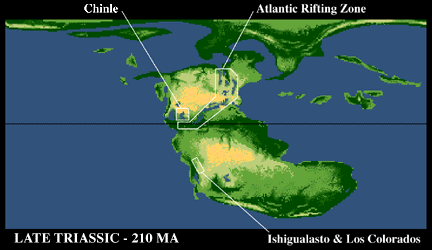
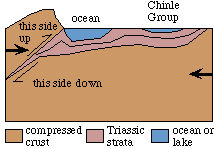
At the same time, the convergence of plates on the other side of North America produced a series of gentle but very broad sags in the crust that filled with a relatively thin sequence of continental strata called the Chinle Group. These broad basins extend from western Texas north to Alberta.
 |
 |
| Triassic Chinle Group outcropping at Petrified Forest National Park. Red cylinders are segments of petrified logs. |
Tectonic context (poorly known) of the Chinle Group. |
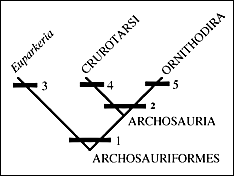
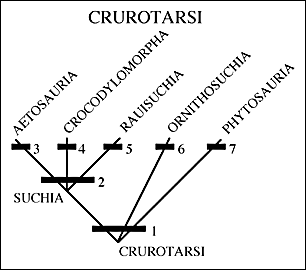
| II. A great many examples of Archosauriformes have been found in the Newark Supergroup and the Chinle Group. To put these forms into context we need to look again at the cladogram (see right) of basic amniote relationships and specifically relationships within the Archosauriformes. | 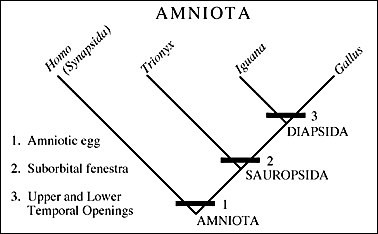 |
 |
As described before, the Archosauriformes have the shared derived character of an antorbital fenestra (see left; 1).
A good representative is Euparkeria who you have seen in previous lectures. Euparkeria lacks specializations that bar it from the ancestry of the archosaurs (3). Almost all of the Archosauriformes from the Newark and Chinle, however, belong to a more advanced sub-group within.The archosaurs have the shared derived character of the loss of teeth on the palate (2). They consist of crocodilians and birds and all of the descendants of that common ancestor (such as dinosaurs in general). |
| Archosaurs can be divided into two groups: the Ornithodira and the Crurotarsi, with shared derived conditions of the ankles being the most important characters. Ankle types within the Archosauriformes fall into three broad categories. First the primitive condition in which the ankle beds at several places in a rather flexible and loose way. The second type is a derived condition called crurotarsal in which the two major bones of the ankle (the astragulus and calcaneum) form a hinge joint (4 above). Usually the calcaneum develops a large backward heel not unlike our own. Crocodiles have this kind of ankle. The third form is another derived form in which a roller-type hinge develops between the astragalus plus the calcaneum and the distal tarsals or metatarsals (5 above). This is called a mesotarsal joint and it is the type seen in birds and other dinosaurs. | 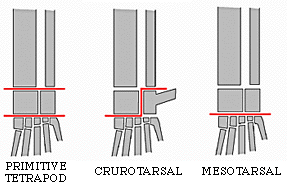 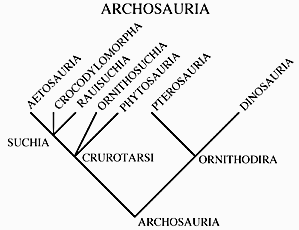 |
The primitive phytosaur Paleorhinus from the Malari Formation of India. Based on Chatterjee (1978).
 |
 |
| Skull of juvenile phytosaur from the Newark basin (Newark Supergroup), North Bergen, NJ. | Skull of phytosaur as found from the Fundy basin (Newark Supergroup), Digby, Nova Scotia, Canada. |
 |
The Phytosauria have the shared derived character of an elongated snout with nostrils that have moved posteriorly (like in a whale) (7). They were very common semiaquatic forms extremely convergent on modern crocodiles in their overall habitus. They can at once be distinguished from crocodiles by the nostrils, which are located towards the rear of the snout, often just in front of the eyes. Rutiodon is a common example from the Newark and the Chinle. They became extinct at the end of the Triassic. |
 |
 |
| Discovery of a phytosaur skull in the Chinle Group, Petrified Forest National Park. Kevin Padian, leader of the expedition is on right. The phytosaur skull is the black rubble they are staring at. | Manus and pes impression of a phytosaur (the track is called Apatopus), from the Newark Supergroup, Newark basin, Oaks, PA. |
Ornithosuchus, based on Walker (1964).
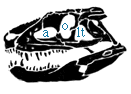 Skull of Postosuchus; after Long and Muray (1996). |
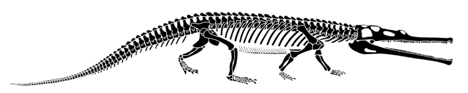
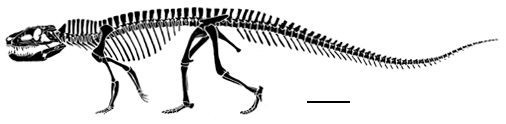
The skull of Postosuchus clearly shows the triangular lower temporal fenestra that is the shared derived character of the Suchia, as well as the slit between the maxilla and premaxilla that is the shared derived character of the Rauisuchia. Abbreviations are - a, antorbital fenestra; lt, lower temporal opening; o, orbit. |
| The foot of Postosuchus clearly has a crurotarsal ankle that can be seen on the photograph to the right. Abbreviations are: c, calcaneum; f, fibula; t, tibia. In this particular individual, the calcaneum and astragalus have fused together because of arthritis. There are also several bony outgrowths on the phalanges that were probably quite painful in life. |
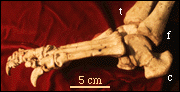 Foot of Postosuchus from North Carolina |
The Aetosauria were herbivorous armored forms. They have a pig-like snout and peg-like teeth. Their shared derived character is the heavy coat of armor covering the back (3). Desmatosuchus from the Chinle and Stegomus from the Newark of New Jersey and Connecticut is a good example.

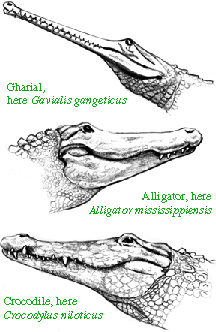 The Crocodylomorpha are by far the most diverse forms and are still with us today. The shared derived character for the group is to have a pair of very elongated carpal bones (the radiale and ulnare [4]). These probably act to add an extra joint to the fore leg increasing its swing. Triassic crocodylomorphs were very graceful, slender forms - more reminiscent of a greyhound than a true crocodile. Some modern crocodiles such as one form from Australia can still move very quickly and can even run in a full gallop [fig]. This was almost certainly true of the Triassic forms. It was not until after the extinction of the phytosaurs at the end of the Triassic that the crocodylomorphs evolved into crocodiles of modern aspect, although fully terrestrial forms persist into the Cretaceous. Some Triassic (and later) forms even evolved into herbivorous forms and forms with very mammal-like teeth.
The Crocodylomorpha are by far the most diverse forms and are still with us today. The shared derived character for the group is to have a pair of very elongated carpal bones (the radiale and ulnare [4]). These probably act to add an extra joint to the fore leg increasing its swing. Triassic crocodylomorphs were very graceful, slender forms - more reminiscent of a greyhound than a true crocodile. Some modern crocodiles such as one form from Australia can still move very quickly and can even run in a full gallop [fig]. This was almost certainly true of the Triassic forms. It was not until after the extinction of the phytosaurs at the end of the Triassic that the crocodylomorphs evolved into crocodiles of modern aspect, although fully terrestrial forms persist into the Cretaceous. Some Triassic (and later) forms even evolved into herbivorous forms and forms with very mammal-like teeth.

Dwarf West African Crocodile in a highwalk mode of locomotion.
| Erpetosuchus originally from Scotland and recently found in the Newark Supergroup of Connecticut is an example of one of these early, terrestrial crocodylomorphs. On the right, "a" is the skull of Erpetosuchus from Connecticut, and "b" is the skull from Scotland (based on Olsen et al., in prep.). Scale is 5 mm. | 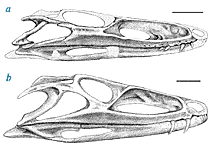 |
IV. Ornithodira
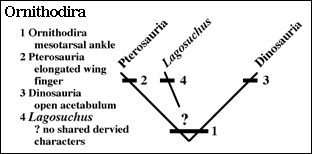 The Ornithodira are the bird-like archosaurs and they include dinosaurs, pterosaurs and incompletely known forms. As stated before their shared derived characteristic is a mesotarsal ankle.
The Ornithodira are the bird-like archosaurs and they include dinosaurs, pterosaurs and incompletely known forms. As stated before their shared derived characteristic is a mesotarsal ankle.
The Pterosauria are the most completely known of the non-dinosaurian ornithodirans to arise in the Triassic. Their earliest appearance is from the Late Triassic of northern Italy, Germany, and Greenland.
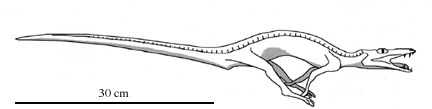
Their shared derived character is the very elongate finger that supports a wing. We will look at them in more detail in the next lecture.Another early ornithodiran is Lagosuchus, known from several partial skeletons from the Ischigualasto Formation. It is a small lightly built animal with very long legs.
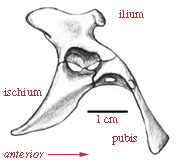 Pelvis of Lagosuchus; after Arcucci (1996). |
The pelvis has an acetabulum (hip socket) with an incomplete rear wall, similar but not identical to the open acetabulum seen in the Dinosauria. Lagosuchus is probably quite similar to the common ancestor of both pterosaurs and dinosaurs. |
Late Triassic units younger than the Ischigualasto Formation have produced much more in the way of remains of dinosaurs. One of the two most notable forms is Coelophysis known from dozens of articulated skeletons from the Chinle Group.
The so-called Whittiker quarry at Ghost Ranch has produced well over 100 articulated skeletons of a small, lightly built dinosaur identified as Coelophysis bauri by Colbert.The genus was originally established by Edward Drinker Cope from very scrappy material collected elsewhere in the Chinle. This original material appears somewhat different from the Ghost Ranch specimens and therefore the name Rioarribasaurus was proposed for the much more complete skeletons. This created a huge uproar among some paleontologists because Coelophysis was such a famous name and had been so long associated with the Ghost Ranch skeletons. The matter was surprisingly acrimonious and ended up being submitted to the International Commission on Zoological Nomenclature (ICZN). Recently, the ICZN ruled that the name Coelophysis should indeed apply to the Ghost Ranch specimens and that a new type specimen should be picked. As a consequence someone will probably end up renaming the original specimens described by Cope!
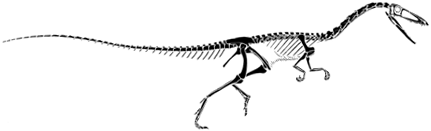
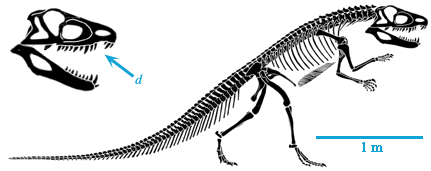
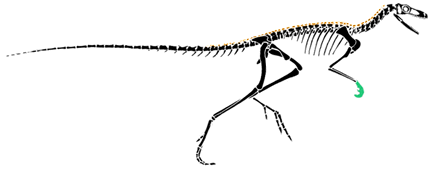
| Coelophysis is a small, very delicate dinosaur characterized by a relatively long neck and an elongate skull. In contrast to Herrerasaurus, the foot is narrow with only three prominent toes. | 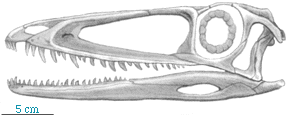 Skull of Coelophysis. After Paul (1988). |
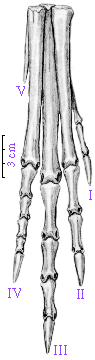 Right pes (foot) of Coelophysis. |
The functionally three toed foot of Coelophysis is a shared derived character of a group called the Theropoda. Theropods make up the bulk of all carnivorous dinosaurs and include such familiar dinosaurs as Tyrannosaurus rex and Velociraptor as well as birds. |
| Coelophysis also has a specialization in the pelvis seen in a number of other theropod dinosaurs. The bones of the pelvis fuse together in the adult, presumably ceasing growth. This is a shared derived character for the Ceratosauria, named after the Late Jurassic genus Ceratosaurus, which we will discuss in a later lecture. Curiously, the best known early ceratosaurs seem to be more specialized than the later ones. | 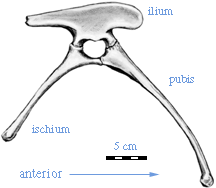 Pelvis of Coelophysis |
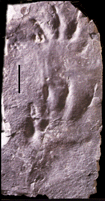
 Grallator parallelum track from Furnace Hill, PA. (track found by Mike Szajna and Brian Hartline) |
Theropod tracks called Grallator and Anchisauripus appear in Newark Supergroup rocks in strata of early Late Triassic age and become larger and more abundant into younger strata. The structure of this kind of track suggests that it was probably made by a small (?young) theropod dinosaur similar to Coelophysis. |
| Present in the same rocks are stubbier footprints, usually with a manus impressed during quadrupedal walking. The manus has tiny claws and thus the tracks cannot have been made by a theropod dinosaur. These tracks are called Atreipus; and they are very common until the Latest Triassic. They most likely belong to an ornithischian dinosaur for which we completely lack skeletal remains. |  Atreipus track from Furnace Hill, PA. (track found by Mike Szajna and Brian Hartline) |
Because we can never be sure exactly what species of trackmaker would make a given kind of track, separate names are given to the tracks. In addition, many species of trackmaker can make the same basic kind of track so that a species of dinosaur track called, say, Grallator parallelum, might have been made by several species or even genera of theropod dinosaur tracks.
Three types of posture can be identified in the Archosauriformes (seen also in the Synapsida independently).
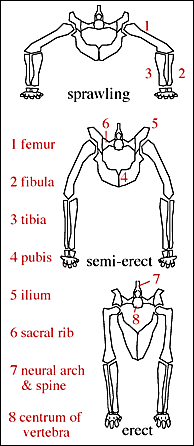 A sprawling gait is primitive for the Amniota and consists of having the limbs in a perpetual "push-up" position. This posture is associated with sinuous motion of the body during locomotion. In this way each limb moves through an arch largely in a plane parallel to the ground. Living lizards have this posture and can move very fast. However, the sinuous motion of the body alternately collapses the lung on each side, preventing breathing during running. That's why lizards have to stop completely and pant after they run, even though they risk capture. A sprawling posture and gait was probably typical for most non-Archosaur Archosauriformes.
A sprawling gait is primitive for the Amniota and consists of having the limbs in a perpetual "push-up" position. This posture is associated with sinuous motion of the body during locomotion. In this way each limb moves through an arch largely in a plane parallel to the ground. Living lizards have this posture and can move very fast. However, the sinuous motion of the body alternately collapses the lung on each side, preventing breathing during running. That's why lizards have to stop completely and pant after they run, even though they risk capture. A sprawling posture and gait was probably typical for most non-Archosaur Archosauriformes.
Crocodiles have a semi erect posture (sometimes called semi-improved). When moving slowly, their posture is sprawling, but when they run, their body straightens out and they pull their legs nearly under their body. When they carry their bodies more nearly erect, they are said to exhibit a "high walk". In this posture the lungs can still operate. Some crocodiles are even capable of galloping. Galloping is the fastest type of longer term quadrupedal motion in which all four feet are off the ground simultaneously during one phase of the step cycle. Most non-ornithodiran Archosauria certainly had the semi-erect posture.
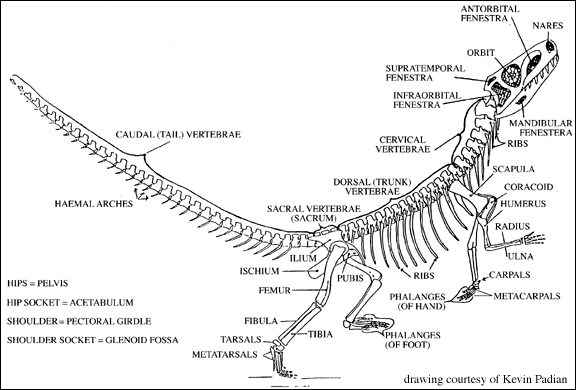
The Ornithodira have an erect posture and gait (often called fully improved) in which the limbs are held directly under the body and flexed in a plane parallel to the body axis. The limbs move in this parasagittal plane during walking slowly as well as running. This allows easy breathing while running. The open acetabulum is the culmination of the permanent attainment of the erect posture.
The open acetabulum is associated with a series of logical mechanical features related to the erect posture and gait of dinosaurs. First, the femur is bent proximally, at right angles. With the femur in the parasagital plane, forces are directed ventrally to the dorsal part of the cup of the acetabulum. In the sprawling and semi-erect postures, forces are directed towards the mid-line of the body, and thus the acetabulum needs to have a bony wall to resist these medially (towards the mid-line) directed forces. In the fully erect posture, dinosaurs had no need to buttress the inside wall of the acetabulum. It could therefore be made of softer (and lighter) tissue.
Perhaps most importantly, the erect posture allows for full-time bipedalism (walking on the hind legs only), thus freeing the hands for grasping. Bipedality seems to have been another shared derived character for dinosaurs, although it was lost in some later groups.
We should note that while during the Late Triassic both mammals and dinosaurs evolved, mammals stayed in the undergrowth (quite literally) for the next 145 million years until the great extinction at the K-T boundary. As Bob Bakker has pointed out, the dinosaurs were probably superior both in physiology and locomotion to mammals until the non-bird dinosaurs were no more. Only then did mammals evolve the erect posture and gait.
Olsen, P. E., Sues, H.-D., and Norell, M., in prep, The Scottish archosaur Erpetosuchus from Connecticut. for American Museum Novitates.
Olsen, P. E. and Baird, D., 1986, The ichnogenus Atreipus and its significance for Triassic Biostratigraphy. In Padian, K. (ed.), The Beginning of the Age of Dinosaurs, Faunal Change Across the Triassic-Jurassic Boundary, Cambridge University Press, New York, p. 61-87.
Paul, G. S., 1988, Predatory Dinosaurs. New York, New York Academy of Sciences, 464 p.
Sereno, P. C., 1993, The pectoral girdle and forelimb of the basal theropod Herrerasaurus ischigualastensis. Journal of Vertebrate Paleontology, v. 13, no. 4, p. 425-450.
{kind=link}