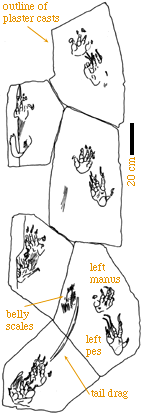
Crude Notes follow.
II. LETS LOOK AT RUNNING - GETTING AT ACTIVITY LEVELS
A. FOOTPRINTS ONLY REAL INDICATION OF HOW DINOSAURS
MOVED AND IT IS REASONABLE TO ASK IF THEY CAN TELL
US ABOUT ACTIVITY
1. POSTURE OBVIOUS FROM TRACKWAYS - BUT ACTIVITY
LEVELS NOT
A. IT IS POSSIBLE TO GET SPEED FROM TRACKS
B. WHEN YOU RUN YOUR PACE AND STRIDE INCREASE
SUGGESTING A WAY
SLIDE OF RUNNING PG 3.7 MCN ALEX
1. SHIP DESIGNERS NEED WAY OF MAKING MODELS
WAY GRAVITY INFLUENCES MOTION IS NOT
PROPORTIONAL TO LENGTH
2. THERE IS A RELATION CALLED DYNAMIC
SIMILARITY WHICH TAKES THIS INTO ACCOUNT
I.E..
3. THIS VALUE TENDS TO BE VARY IN PROPORTION
FOR DIFFERENT SIZE SHIPS
4. R. McNEILL ALEXANDER - HAS WORKED OUT A
SIMILAR RELATIONSHIP FOR RUNNING TETRAPODS
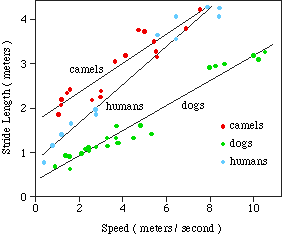
RELATIONSHIP BETWEEN STRIDE AND SPEED
5. SUGGESTED RELATIONSHIP
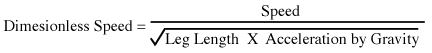
NOW WE CAN GRAPH DIMENSIONLESS SPEED AGAINST RELATIVE STRIDE, AS FOLLOWS
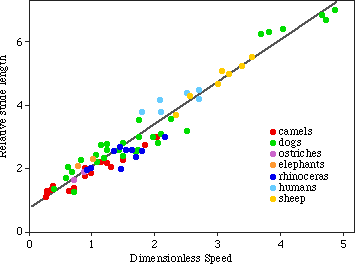
6. WE CAN SOLVE FOR SPEED GIVEN THIS RELATIONSHIP
7. BUT WE STILL HAVE TO GET LEG LENGTH FROM A FOOTPRINT
8. WE DO THIS BY OBSERVING THAT LEG LENGTH SCALES
TO FOOT LENGTH FOR DINOSAURS NO MATTER WHAT THE
SIZE OR WHETHER THEY ARE BIPEDAL OR QUADRUPEDAL
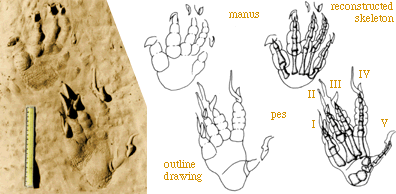
SLIDE ALBERTOSAURUS A TYRANNOSAUR
SLIDE DATA ON LIMB VS FOOT
IN FACT LEG = 4 X FOOT (to metatarsals)
9. THUS, FOR A VARIETY OF DINOSAUR TRACKS WE GET
THE FOLLOWING RELATIONSHIPS
SLIDE OF TABLE OF SPEEDS
MOST DINOSAURS TRACKS ARE THUS OF WALKING ANIMALS
BUT SOME COULD RUN FAST, AS FAST AS THE FASTEST MAMMALS
SO THIS SHOWS DINOSAURS COULD MOVE AS FAST AS FASTEST MAMMALS
II. GETTING AT METABOLISM FROM DINOSAURIAN
STRUCTURAL PARALLELS
A. NOW WE CAN SEE THAT DINOSAURS COULD PROBABLY
RUN VERY FAST BUT SO CAN SOME REPTILE FOR
SHORT DISTANCES
B. STRUCTURALLY, HOWEVER, THEY ARE MAMMAL OR BIRD LIKE
1. STRUTHIOMIMUS - OSTRICH
2. DEINONYCHUS - ARCHAEOPTERYX
3. HADROSAUR - ELK
C. LARGE ANIMALS HAVE SIMILAR STRCTURAL REQUIREMENTS
1. SAUROPOD AND ELEPHANT
III. INTERNAL CONSTANCY VS VARIABILITY OF TEMP
A. WHAT ABOUT HEAT PRODUCTION
1. RESTING METABOLIC RATE GREATER IN MAMMALS
AND BIRDS THAN IN REPTILES
SLIDE OF RESTING METABOLIC RATE FROM MC A
2. SIZE CONFERS SOME THERMAL INERTIA
1. SURFACE AREA IS WHERE HEAT IS LOST
2. SURFACE AREA INCREASES AS THE SQUARE OF linear DIM
3. BUT VOLUME INCREASES AS THE CUBE
4. THEREFORE FOR THINGS OF SIMILAR SHAPE,
LARGE THINGS LOSE HEAT SLOWER
5. BIG ANIMALS HAVE GREAT THERMAL INERTIA
SO BIG DINOS WERE COULD HAVE BEEN INERTIAL
HOMEOTHERMS
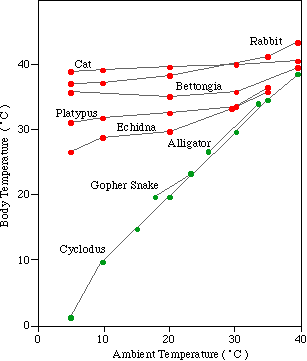
SLIDE DATA ON LITTLE VS BIG TEMPS
3. ALSO THIS MEANS THAT LARGE THINGS WITH
HIGH METABOLIC RATES NEED TO DUMP HEAT
4. BUT SMALL THINGS LIKE SHREWS NEED TO
CONSERVE IT ALWAYS
5. SOMETIMES BIRDS WITH GOOD INSULATION NEED
TO COOL - AIR SAC SYSTEM
6. AIR SAC SYSTEM PRESENT IN GIANT SAUROPODS
- PROBABLY FOR COOLING
B. OF COURSE, IF YOU HAVE HIGH METABOLIC RATES
YOU NEED TO EAT MORE FOOD
SLIDE OF BAKKERS DOG VS LIZARD PG 376
1. THUS WE WOULD EXPECT THE SAME NUMBER OF PREY
ANIMALS TO BE ABLE TO FEED A LARGE NUMBER OF
LOW METABOLIC RATE ANIMALS THAN HIGH ONES
2. IN FACT IF WE LOOK AT THE DISTRIBUTION OF
PREDATOR-PREY RATIOS WE CAN SEE THAT DINOSAURS
HAVE A LOWER PREDATORY/PREY RATIO THAN EOCENE MAMMALS
SLIDE OF PREDATOR/PREY RATIOS DRAWING FROM BAK PG 351
3. THUS IT EVIDENTLY TOOK MANY, MANY HERBIVOROUS
DINOSAURS TO FEED ONE CARNIVOROUS ONES
IV. ACTIVITY LEVEL VS. BONE STRENGTH
1. IT HAS LONG BEEN OBSERVED THAT THE STRUCTURE
OF THE BONES IN MAMMALS AND BIRDS IS DIFFERENT
THAN THAT SEEN IN OTHER LIVING REPTILES
ADAPTED SLIDE FROM BAKKER PG 352
HAVERSIAN SYSTEM PRESENT IN BIRDS AND MAMMALS
EXPLAIN
SLIDE OF HAVERSIAN SYSTEM AND NOT
EXPLAIN CROMPTON'S ARGUMENT
V. PHYLOGENETICS
SHARED DERIVED CHARACTERS OF DINOSAURS AND BIRDS
FOUR CHAMBERED VS THREE CHAMBERED HEART
VII. SUMMARY
A. DINOSAURS COULD RUN VERY FAST
B. DINOSAURS WERE BUILT FOR LONG PERIODS OF ACTIVITY
C. LARGE DINOSAURS COULD HAVE EASILY BEEN INERTIAL
HOMEOTHERMS BUT SMALL ONES HAD TO PRODUCE EXTRA
HEAT OR WOULD HAVE TO ADAPT THEIR BEHAVIOR TO
RECEIVE IT FROM ENVIRONMENT
D. LARGE DINOSAURS NEEDED TO LOSE HEAT
E. PREDATOR/PREY RATIOS SUGGEST THAT CARNIVOROUS
DINOSAURS NEEDED AS MUCH FOOD AS CARNIVOROUS MAMMALS
F. DINOSAUR BONES REMODELED AS OFTEN AS MAMMAL AND
BIRD BONE SUGGESTING HIGH ACTIVITY LEVELS
G. BIRD ARE DINOSAURS AND MAY HAVE INHERITED THEIR
METABOLIC SYSTEMS
CONCLUSION:
AT LEAST SOME DINOSAURS WERE AS ACTIVE AND
"WARM BLOODED" AS ANY MAMMAL OR BIRD
AND SO ALTHOUGH WE CANT GO BACK IN TIME AND
MEASURE THEM WE CAN INFER, MAKE PREDICTION AND
TEST THEM IN THE FOSSIL RECORD
NOTE THAT COMBINED WITH THEIR LOCOMOTOR ADAPTATIONS
IT BECOMES EASY TO SEE WHY DINOSAURS REMAINED
DOMINANT FOR SO LONG OVER MAMMALS
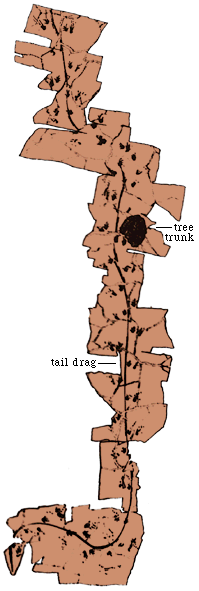
Dinosaur Sociality from Tracks