All during this time the earth was still being bombarded by asteroids and comets, a process that still occurs but at a much lower (although still significant!) rate.
At some point however, the Moon formed. Exactly how that happened is a major subject of debate. One theory claims that the Moon is a tiny planet captured by the Earth's gravity. The other is that the moon was literally splashed out of the Earth by the impact of a Mars-sized planet. The latter theory is favored now because it explains some odd but important features of the Earth's and Moon's chemical composition.
The Earth was reborn during the iron catastrophe and maybe again by the formation of the Moon. Any trace of surface structure was wiped out by the melting.
IV. However, the crust finally solidified by about 3.7 billion years ago. Gasses pouring out of volcanoes and fissures, along with lava, began to accumulate, perhaps added to by the impact of a few giant comets (which are mostly gas).The gases that accumulated were those we still find coming out of volcanoes:Water vapor (H2O)
Hydrogen chloride (HCl)
Carbon Monoxide (CO)
Carbon Dioxide (CO2)
Nitrogen (N2)
These gases combined to form:Methane (CH4)
Ammonia (NH4)
Hydrogen Cyanide (HCN)
This atmosphere would be quickly fatal to us.
V. As the crust cooled water would condense and accumulate as oceans. This happened very soon after the crust solidified.
VI. LIFE EVOLVES
The origin of life is shrouded in mystery. But there have been significant steps towards some understanding.
Making many complex organic molecules is the first step, but apparently not a difficult one.
This was shown in the famous Miller-Urey experiments done in the 1950's.
Stanley Miller and Harold Urey were a graduate student/professor team here at Columbia. They planned to set up an experiment to see how complex organic molecules could be produced. Basically they mixed together gasses that they thought the primitive Earth would have in a jar and zapped the gases with an electrical spark. Voila! Complex organic goo collected at the bottom of the jar. Included were amino acids, the building blocks of proteins. But no life.
Such experiments have been repeated many times and it is clear that it is easy to make many complex organic compounds but none of these simple experiments produced even the basics of life.
The two essential elements of Life are that the system needs to be able to replicate and it needs to be separated from its surroundings.
What is still pretty much a mystery is the origin of heredity and reproduction. It's possible that the surface of clay or some other mineral acted as a template for organization, but this is very hypothetical, and key experiments have yet to be performed or even rationalized.
Here is a link to a some new experimental (and highly technical) work on creating replication.Simple self-replicating molecules developed in this so-called primordial soup of organic matter in water.
But once there was replication, there would be occasional errors and hence, there could be evolution.
B. However it happened, we are fairly sure it happened early in Earth's history because fossil carbon in 3.7 billion year old rocks and stromatolites from about the same time have the tell-tale signs of life.
By 3.5 billion years ago there was life, certainly with the full compliment of the basic genetic system - with DNA and RNA. (Click here if you want a basic outline of DNA and RNA) (Click here if you want lots of detail)
These were cells encapsulated in membranes so their internal workings were isolated from their surroundings.
We are positive that the earliest life forms were prokaryotes, such as bacteria and Cyanobacteria.
Evidently, photosynthesis must have started nearly at the beginning. At once that changed the world because of the release of free oxygen.
Evidence for this are stromatolites and silicified microorganisms.
At first all sediments were gray, indicating reduced iron. Great gold deposits formed such as the Witswatersrand in river deposits associated with iron sulfides. This is not possible in an O2-rich atmosphere.
C. Up to 2 billion years ago the O2 produced by photosynthesis was used up by the oxidation of reduced iron, Fe2+ to Fe3+. Oxygen, an oxidizing agent, is an electron acceptor. Iron is soluble in compounds in the Fe2+ state but insoluble in Fe3+ compounds such as Fe2O3.
Then about 2.6 billion years ago we get the first red Banded Iron Formations (BIF's) - red and gray zones of oxidized iron layers of silica.
Responsible for world's most important iron deposits.
Cycles in photosynthesis thus produced cycles in O2 in the water column, which produced cycles in the oxidation and then deposition of Fe3+ compounds on the ocean floor. Likewise the drawdown of CO2 in the water column would produce an increase in the pH of the water and an increase in the solubility of Si. But when photosynthesis was operating at lower levels, the pH went down and the deposition of Si would take place.
Formation probably due to cycles of photosynthesis then biological crash. Soluble iron comes out of solution in presence of O2 when photosynthetic microbes doing well. When they (perhaps seasonally) died off, the silica was deposited.
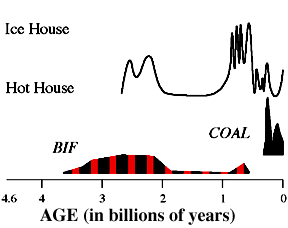 |
As soluble iron started to get used up, O2 began to build up in atmosphere, and CO2 dropped.
This continues until 2 billion years ago, when we see the first appearance of red beds.
Red beds are very shallow-water, river, or soil deposits in which the iron has combined with O2 to form red iron oxide. |
These red beds indicate that O2 is building up in the atmosphere.
About the same time, BIFs decline, an indication that reducing compounds are disappearing from the oceans.
The basic set-up of weathering as we know it was probably set up at this time.
Experiments show that even algal scum on crushed rock enhances weathering by several times to several orders of magnitude.
VII. UNTIL ABOUT 1.9 BILLION YEARS AGO WE HAVE EVIDENCE ONLY OF THE SIMPLEST KINDS OF LIFE - THE PROKARYOTES - BACTERIA AND BLUE GREEN ALGAE
But at about 1.9 billion years we start to see fossils of much larger cells. These cells belong to the Eukaryotes - of which we are members.
Eukaryotic cells are symbiotic colonies of prokaryotes; many of the symbionts are called organelles. O2 is handled by Mitochondria, chloroplasts handle photosynthesis. These organelles independently replicate with their own genome DNA sequences in circular strands, as in bacteria. Several other kinds of organelles are similar.
The relatively independent lives and separate genetic systems of these organelles led Lynn Margulis to develop the "organelle theory" (a form of which was first proposed by Mereschkowsky in 1905) in which different groups of prokaryotes became endosymbionts in other cells.
Eukaryotic metabolism can get going at about 2 % of present O2 levels, which is consistent with the appearance of common red beds at about the same time. Some photosynthetic eukaryotic organisms developed colonial forms and the first "sea weeds occur". This is the first good evidence of eukaryotic multicellularity.
The development of sex allows for species in the same sense we know them know lineages of interbreeding individuals.
About 650 million years ago (0.65 billion) the first apparently multicellular forms are present. These are called the Ediacara assemblages. They all seem to be elaborations of forms with large surface areas, all living in shallow relatively high energy environments - often in red beds.
Perhaps they needed greater surface area to absorb O2 in less-than-modern atmospheric amounts.
Although superficially similar to several animal phyla, it is unclear if ediacarans belong to extant groups. Adolph Seilacher places them all in their own phylum, which he calls the Vendizoa.
The key to multicellular animals seems to the so-called homeotic genes. They are very odd and newly discovered. Seem to be involved with headness, tailness, very general properties. So head structures are homologous between flies and humans. So are eyes, even though a fly's eye is compound and ours are not, and our common ancestor had no eyes at all. The instructions for eyeness is evidently homologous. This sophistication is probably why multicellularity took so long.
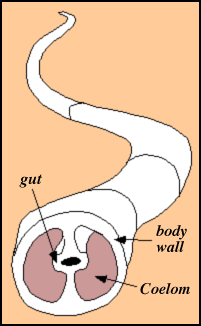
About the same time (650 million years ago) the first burrows appear.
This records the evolution of the coelom: a sac that contains the organs. We have one. It allows for hydrostatic support of the body so it can push through mud, etc.
This allows larger organisms that can have a hydrostatic skeleton and the ability to bend and twist and push even though they are large. Major modification of the sediments result. And oxidative processes can now go on at depth. Hence the efficiency of use of carbon fixed by photosynthesis increases.
The basic structure of a coelom.
Now animals and plants could modify the physical structure of their environment, not just its chemistry - and eat each other.
E. In the Proterozoic, the Earth's climate seems to have gone through great swings. There may have been glaciers in the tropics at times, alternating with times of more normal carbonate deposition as we see today in the warm regions. This suggests oscillating CO2 levels. I suspect that CO2 levels were basically at the whim of tectonic events during this period: undamped forcing with few damping negative feedbacks resulting in dramatic oscillations. This is a major focus of research.
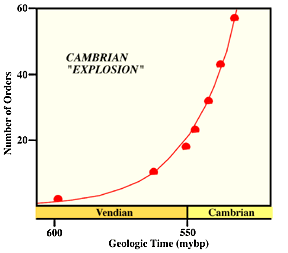 |
VIII. At the beginning of the Cambrian 540 million years ago multicellular animal life explodes!
There is a massive increase in the kinds of multicellular animal present. Could be due to the first wave of multicellular plants on land - perhaps lichens - but very poor evidence. But that would increase the burial rate of organic carbon which would allow for many more animals in the oceans. |
This is the Cambrian explosion. A famous example is the Burgess Shale from British Columbia. It looks like most major animal phyla, perhaps all of them, were around by the middle Cambrian. By the end of the Cambrian the Phanerozoic pattern of marine organisms was pretty much established.
Evolution of taxa is very rapid - looks exponential.
IX. EVOLUTION OF TAXONOMIC DIVERSITY DURING THE REST OF THE PALEOZOIC HAS A VERY INTERESTING PATTERN.
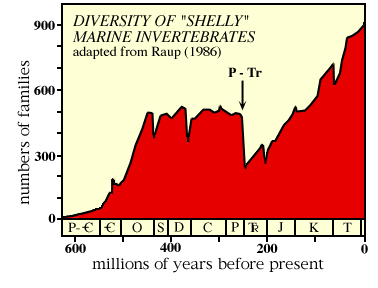
The above figure shows diversity of families of shelly marine animals during the Phanerozoic. P - Tr marks the end-Paleozoic (=end-Permian) mass extinction.
Basically, it looks like after an explosive start, the diversity of life pretty much leveled out, with a few downward blips (actually mass extinctions).
X. The next major event was the development of vascular land plants, beginning in the Ordovician and culminating in the Late Devonian. Land plants became bigger and bigger and more and more divorced from relying on water for reproduction.
Ferns and their allies reproduce with alternation of generation. The fern you see is the so-called sporophyte generation. It produces spores which float though the air, land and sprout into green liver-like gametophytes, which shed egg and sperm into water which unite with eggs to produce the next sporophyte generation.
They need water to reproduce, which is why they are most abundant in wet areas.
By the late Devonian the seed was invented. Here the sporophyte plant produces a male gametophyte which becomes a pollen grain, which floats in the air to the female gametophyte, still attached to a sporophyte. The male gametophyte puts down a pollen tube, fertilizing the egg in the female gametophyte. An embryo develops with a storage of food in a hard capsule, which we call a seed.
No water needed for this reproduction. Trade off is that dispersal is slower since the seed is so very much bigger.
Now the locus of the Earth's biomass shifts from the sea to the land.
By the late Devonian there is a fairly diverse soil community, very similar to what we will see when we look at modern soils later on. Mites, spring tails, millipedes and lots of carnivores to feed on them.
XI. But vascular plant material is tough stuff. Once buried the lignin in woody material hardly breaks down at all. Early on in the Devonian and in the next period, the Carboniferous, there is hardly anybody big around to eat the woody matter.
Some sucking insects, a very few leaf biters, even rarer wood burrowers.
In the late Devonian amphibians evolve and by the middle Carboniferous we have the origin of the Amniotic egg, which frees terrestrial vertebrates from the water in the same way the seed does for plants.
But even towards the end of the Carboniferous, there are no full-time vertebrate herbivores - just a few with teeth that could handle some leafy or fruity tissue. In fact the terrestrial vertebrate community was dominated by carnivores - with the base of the food chain being aquatic, not land plant based. If you look at just the land animals the food pyramid looks upside down.
It is during this time that the rate of burial of terrestrial plant matter dramatically increases as seen in the delta 13C curve. Some say this is due to the development of vast swamps. In swamps, the decay of vascular plant matter is impeded by burial in water-saturated conditions, preventing aerobic decay.
But I believe this is due to the lack of abundant herbivory which now mills down vascular plant matter to high-surface-area bits that can be readily attacked by bacteria, fungi, and small soil animals.
Hence the ecosystem efficiency of the terrestrial world was low and carbon accumulated in a much wider range of environments than now.
As a consequence of the burial of this organic matter, O2 levels probably reached our current levels in the Carboniferous, but were kept from going higher by fire and the burial of nutrients.
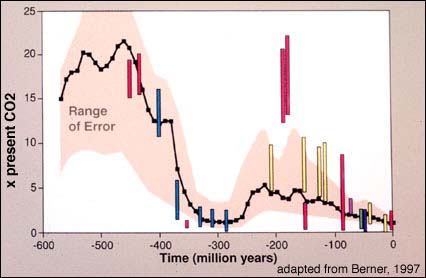
The above figure shows Phanerozoic CO2 levels. The curve represents the results of a geochemical model of CO2 and the bars are the range of paleo-CO2 measurements based on soil calcium carbonate.
The plants also fertilized the chemical weathering process which almost certainly massively increased the burial rate of inorganic carbon as carbonate. The enlargement of the pool of carbonate in the oceans kept the equilibrium levels of CO2 in the atmosphere much lower.
XII. During the peak of land plant spread, in the Late Carboniferous (300 million years ago), the world plunged into a long glacial age, almost certainly due to plant-mediated weathering.
Perhaps we came out of this glacial world in the Permian because of the evolution of many new herbivores. Vertebrate herbivores finally become more abundant than carnivores and for the first time the familiar ecological pyramid is recognizable.
A major driving force for this other, than the terrestrial vertebrates, were certainly the insects and termites which may well have evolved in the Permian.
Ecosystem efficiency rose, the burial of plant matter slowed, and chemical weathering slowed.
We moved from the ice house to the greenhouse.
XIII. The end of the Permian saw the greatest mass extinction of all time. Raup has estimated that perhaps 99% of all species went extinct.
The origin of this mass extinction is unknown, however, there is speculation that massive amounts of CO2 and sulfuric acid released by the massive volcanism that produced the Siberian Traps (basalts) may have contributed by causing sever climatic disruption.
Whatever the origin of the great end-Permian mass extinction, the following period ushered in the Age of Dinosaurs.
Go to Lecture 8
BACK TO SYLLABUS
BACK TO THE DINOSAUR AND THE HISTORY OF LIFE HOME PAGE


