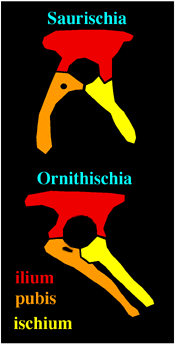
 The Ornithischia have the shared derived character of a backwardly pointing pubis.
The Ornithischia have the shared derived character of a backwardly pointing pubis.
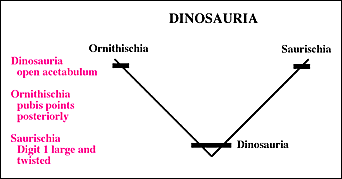 The Saurischia comprise the dinosaurs with the shared derived character of a hand in which digit 1 is enlarged and twisted so that the hand becomes one that can grasp. The Saurischia are the so-called "lizard-hipped" dinosaurs because most saurischian groups retain the forward pointing pubis seen in more primitive archosaurian. This is called the "triradiate pelvis"
The Saurischia comprise the dinosaurs with the shared derived character of a hand in which digit 1 is enlarged and twisted so that the hand becomes one that can grasp. The Saurischia are the so-called "lizard-hipped" dinosaurs because most saurischian groups retain the forward pointing pubis seen in more primitive archosaurian. This is called the "triradiate pelvis"
The Ornithischia have the shared derived character of a backwardly pointing pubis.
The change in pubis orientation allows for a bigger gut, needed to digest plant material, in the Ornithischia. The center of gravity thus remains at the hips so the animal did not tip forwards.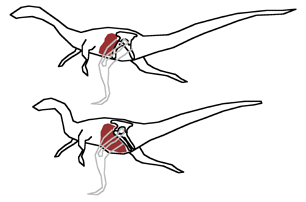
The earliest known ornithischian is the poorly known genus Pisanosaurus from the Ischigualasto Formation of Argentina.
Pisanosaurus is known from only scraps of the pelvis, some limb boneds and toes, and a partial lower jaw and upper jaw (dentary and maxilla, respectively. Other than Pisanosaurus, only scraps of bone and isolated teeth of other ornithischians are known from the Triassic. Footprints, are rather common however, and show that although dinosaurs were not as a rule the dominant members of Triassic communities, ornithischians were an important component, even early on.
 On the right is a photograph of a track called Atreipus, in this case from the Newark basin of Pennsylvania (part of the Atlantic rift system). Note its three-toed front foot. The other two toes were carried off the ground.
On the right is a photograph of a track called Atreipus, in this case from the Newark basin of Pennsylvania (part of the Atlantic rift system). Note its three-toed front foot. The other two toes were carried off the ground.
The reconstruction of Atreipus, seen below, is based on reconstructing the skeleton of the foot in the footprint and than comparing it to what we know of real skeletons. We will talk more about this later on.
We will deal more with ornithischian when we begin to talk about the Jurassic, after the midterm.
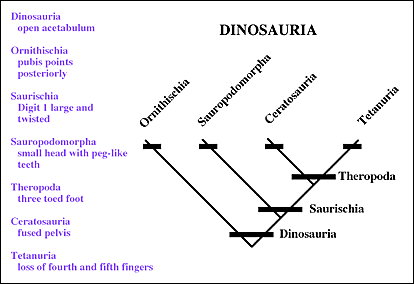
The Sauropodomorpha include the giant long-necked herbivorous dinosaurs such as Apatosaurus (A.K.A. Brontosaurus). The shared derived character of the Sauropodomorpha is to have a small head with peg-like teeth.
The Theropoda include all of the meat-eating saurischians, such as Tyrannosaurus rex as well as birds. Their shared derived characteristic is to have a functionally tridactyl (three toed) hind foot. A small fourth is usually present. It is this sort of foot that is inherited by birds.
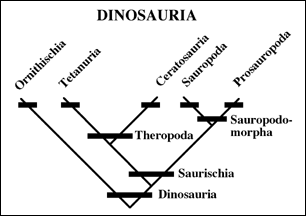 The Sauropodomorpha is split into two groups: the Sauropoda and the Prosauropoda.
The Sauropodomorpha is split into two groups: the Sauropoda and the Prosauropoda.
The Sauropoda have the shared derived character of a forwardly expanded ilium. They do not appear until the Early Jurassic and we will, thus, consider them after the midterm.
The Prosauropoda have the shared derived character of an apron-shaped pubis. This broad surface probably acted as a shelf for the gut when the animals reared up to eat the branches of trees. The best known prosauropod is Plateosaurus, known from dozens of articulated skeletons from the Late Triassic of Germany. Prosauropods, by the way were the first animals to be able to eat a tree.
A famous tiny prosauropod is Musasaurus from Patagonia (no, a dinosaur, not a parka). It is based on a minute skeleton, almost certainly a baby of a Plateosaurus-like prosauropod.
The earliest known purported theropod is Herrerasaurus also from the Ischigualasto Formation of Argentina. In fact, Herrerasaurus does not have a typical theropod foot and it is assigned to the Theropoda mostly on the basis of other skeletal characters by Paul Sereno.
 The Ceratosauria are among the earliest definitive theropod dinosaurs . Their shared derived character is to have a fused pelvis in the adult. The most famous Triassic ceratosaurian dinosaur is Coelophysis from the Late Triassic. It is known from hundreds of skeletons from the Ghost Ranch in New Mexico. There appear to be two sexes (as we might expect) represented in the mass-kill from Ghost Ranch: a robust for and a gracile form. In addition several specimens have babies in their gut which they evidently ate. Mothers may have done this to recoup their investment in their offspring, much as mice do when conditions deteriorate. It is also possible that males may have eaten the offspring of rival males.
The Ceratosauria are among the earliest definitive theropod dinosaurs . Their shared derived character is to have a fused pelvis in the adult. The most famous Triassic ceratosaurian dinosaur is Coelophysis from the Late Triassic. It is known from hundreds of skeletons from the Ghost Ranch in New Mexico. There appear to be two sexes (as we might expect) represented in the mass-kill from Ghost Ranch: a robust for and a gracile form. In addition several specimens have babies in their gut which they evidently ate. Mothers may have done this to recoup their investment in their offspring, much as mice do when conditions deteriorate. It is also possible that males may have eaten the offspring of rival males.
It would be interesting to see whether only robust or gracile forms had babies inside.
There is considerable debate about the name Coelophysis. The scrappy type material of the genus was described by Cope from a different locality than Ghost Ranch. There also seem to be some differences between the type material and the much better preserved Ghost Ranch mass-kill specimens. For that reason the name Rioarribasaurus has been given to the Ghost Ranch material. It is too soon to tell if this proposal will be accepted by the scientific community.
As you will recall, the Synapsida split off from the Sauropsida way back in the Carboniferous near the origin of the Amniota. Early synapsids included the Early Permian sail-backed reptiles Dimetrodon and Edaphosaurus. Dimetrodon was a large carnivore and Edaphosaurus a large herbivore; both living at the same time and the same place. Each evolved their sails independently (they are analogous characters). The sails were almost certainly from regulating their temperature. They probably turned their sails to face the sun when they wanted to gain heat and parallel to the wind to lose heat. Remember, the Late Carboniferous was an "ice-house" world. The carnivore Dimetrodon probably evolved the sail to get a head start on chilly mornings on its prey, and the herbivore (prey) evolved the sail to get away from Dimetrodon! Early on-thermoregulation seems to have been a synapsid concern.
During the Triassic we see a number of changes n the clades that eventually would include mammals. In particular, there is a shift in the structure of the jaw articulation that involved moving two jaw bones from the jaw joint into the middle ear. The condition in which the middle ear has three bones linking the ear drum to the middle ear is what defines mammals. The transition occurred sometimes in the Late Triassic. In addition mammals evolved a tooth replacement pattern with only one set of replacement teeth in which the molars have double roots and the teeth occlude precisely.
So dinosaurs and mammals evolved more or less at the same time. As well preserved mammals are known only from the Jurassic we will discuss this key transition in more detail later.
Plants
The Late Triassic saw a rise in a group of seed-bearing conifers called the Cheirolepidaceae or "cheiroleps". These plants ranged from shrubs to large trees. They tended to have fleshy leaves and may well have been deciduous. Their rise coincides with the rise to prominence of the sauropodomorphs: first the prosauropods of the Triassic and Early Jurassic and than the giant sauropods, later in the Jurassic and Cretaceous. As the cheiroleps became less common though the Cretaceous, so did sauropods.
The sudden drop in animal diversity corresponds to a dramatic change in the plants, with diverse plant community giving way to very low diversity community almost entirely dominated by the cheiroleps (at least in the tropics) in the earliest Jurassic.
A very thin layer has been found in the Newark basin in Pennsylvania that has almost nothing but fern spores in it. Ferns often the first colonizers after a catastrophes and the appearance of this so called "fern spike" in the fossil pollen and spore record appears to be an indication of a major ecological disaster. Abundant reptile footprints in the same area show a massive change in taxonomic composition right at the same layer.
In Austria and Italy what is probably the same layer has grains of a type of quartz called "shocked quartz". Shocked quartz is known from only known asteroid impact sites and nuclear bomb test sites. It is produced by very high pressures, but short lived pressures. This suggests that the Triassic-Jurassic boundary was caused by the impact of a giant asteroid similar to the one that terminated the Cretaceous Period. But, in this case rather than wipe out the dinosaurs the Triassic-Jurassic mass extinction may have opened the doors for the dinosaurs by wiping out their competition.
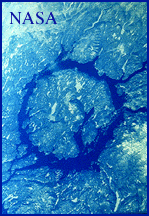 A candidate for an impact event that could be the cause of the Triassic-Jurassic mass extinctions is the great Manicouagan impact structure of northern Quebec, Canada. Thus gigantic structure formed about 200 million years ago by the impact of a ~10 km diameter asteroid. The Manicouagan structure, produced by this impact is easily visible from space and consists of a ring of shattered rock 70 km in diameter surrounding a disk of rock that was completely melted by the impact event. About 1 km of rock has been eroded from the site since the impact and its original size may have been about 100 km in diameter. The amount of energy released by this impact was probably about 10,000 times what the combined energy that would be released by the simultaneous explosion of the entire nuclear arsenals of the United States and former Soviet Union!
Although a possible candidate for the cause of the end-Triassic mass extinction, the newest dates suggest that the Manicouagan impact structure could be about 12-15 million years older than the mass extinction itself, thus ruling it out as a causal agent. We will come back to the evidence for this in later lectures.
A candidate for an impact event that could be the cause of the Triassic-Jurassic mass extinctions is the great Manicouagan impact structure of northern Quebec, Canada. Thus gigantic structure formed about 200 million years ago by the impact of a ~10 km diameter asteroid. The Manicouagan structure, produced by this impact is easily visible from space and consists of a ring of shattered rock 70 km in diameter surrounding a disk of rock that was completely melted by the impact event. About 1 km of rock has been eroded from the site since the impact and its original size may have been about 100 km in diameter. The amount of energy released by this impact was probably about 10,000 times what the combined energy that would be released by the simultaneous explosion of the entire nuclear arsenals of the United States and former Soviet Union!
Although a possible candidate for the cause of the end-Triassic mass extinction, the newest dates suggest that the Manicouagan impact structure could be about 12-15 million years older than the mass extinction itself, thus ruling it out as a causal agent. We will come back to the evidence for this in later lectures.