|
 |
| Latimeria | Protopterus |
BACK TO DINOSAURS 2001 HOME PAGE
During the Devonian, the Tetrapoda arose. These consist of the four legged animals. They evolved from Sarcopterygian fish, living members of which include the coelacanth Latimeria (also see this www page) and the lungfish Protopterus (Africa), Lepidosiren (South America) and Neoceratodus (Australia).
|
|
| Latimeria | Protopterus |
Cladistically, of course, tetrapods and us are in fact fish, but I will use quotes to denote informal use of a familiar, although paraphyletic, group.
Early tetrapods were, like early land plants, tied to the water by their reproductive mechanisms. Like most "fish" they had external fertilization in water and laid eggs in water which developed into aquatic larvae. The larvae metamorphosed into land-living adults. Living "amphibians" have inherited this primitive mode of reproduction.
Sometime during the Carboniferous, one group of tetrapods developed the amniotic egg. Primitively, the amniotic egg has a shell hardened by calcium carbonate which is impermeable to water but allows gases to be transpired. The embryo lies floating in the amniotic fluid, which is formed by the embryo. The shell is formed by the mother.
The amniotic egg is a major shared derived character for the taxa in the Amniota, which includes us as well as the dinosaurs. Soon after the amniotes appeared, two distinct group are recognizable - these are the Synapsida (which includes us) and the Sauropsida (which includes the dinosaurs).
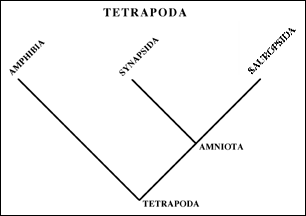
Lets take a look at the Tetrapoda and its major subgroups.
The major groups and their synapomorphies are:
Tetrapoda - 4 legsNow lets look in more detail at the Amniota.
Amniota - Amniotic egg
Lissamphibia (living "amphibians") - Teeth on pedestals
Synapsida - Lower temporal opening (see below)
Sauropsida - Suborbital fenestra (see below)
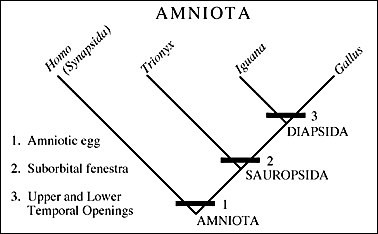
In this case we have replaced the higher level taxon with a representative lower level one. So, the representative synapsid becomes Homo sapiens, and we have replaced the Sauropsida with three taxa. These three taxa are the turtles (= Chelonia) represented by the genus Trionyx, the lizards (= Squamata) represented by Iguana, and the dinosaurs (= Dinosauria), represented by the chicken genus Gallus.
II. Critical data for the relationships within the Amniota is present in the nature of various large openings in the skull called fenestrae (literally windows - fenestra, singular).
We will be using these sorts of characters through the rest of the term.
The complete set of openings is seen in a so-called diapsid - type skull characterized by having two lateral temporal openings. The diapsid-type skull shown here is also an archosaur. A dinosaur might have such a skull.
Others kinds of opening configurations are called anapsid, synapsid, and euryapsid depending on the number and position of the openings on the side of the head, behind the eye.
The names anapsid, synapsid, diapsid, and euryapsid are names of the characters and they do not necessarily characterize monophyletic groups.
Most often they are to increase the area and improve the alignment or the attachment of major muscles.
They also serve to allow an area that expands outward for muscles that would otherwise be in a confined space.
They lighten the skull without compromising strength.
They occasionally have a gland in them, although this is unusual.
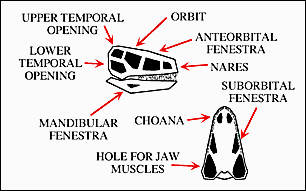
The view of the skull in the lower right of this diagram is looking up from underneath with the lower jaw removed.
A special set of openings are the choanae which are the internal nostrils. In an inhalation, air goes into the nostrils and though the choanae into the mouth cavity and then into the wind pipe (trachea).
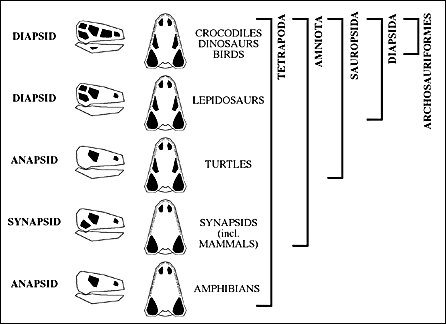
Taken together the different skull openings can be used as shared derived characters that are present in major groups of tetrapods.
The number and arrangement of the fenestra and choanae can be mapped directly onto the cladogram for the Tetrapoda
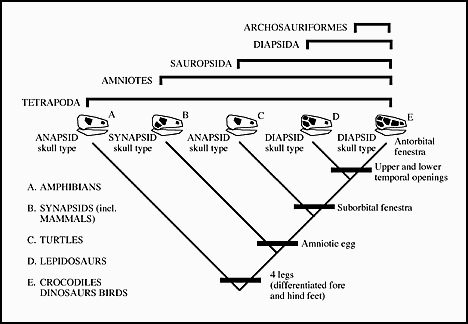
Cladogram showing the different arrangements of skull openings. (note that amphibians here are the Lissamphibia which are a monophyletic group, rather than the paraphyletic "amphibia".
The Diapsida is in turn broken into two main groups.
1. Archosauriformes - antorbital fenestra
2. Archosauria - loss of palatal teeth
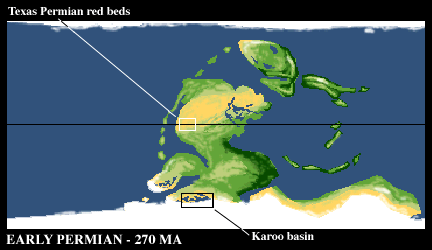
Two famous areas for the Permian are the Early Permian strata of Texas and Oklahoma and the Late Permian strata of the Karoo basin of southern Africa.
As in the later part of the Carboniferous, great glaciers covered the south pole and extended quite far north.
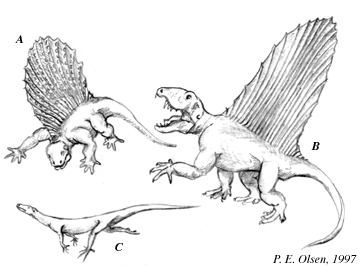
Early Permian terrestrial vertebrates were dominated by synapsids, the most notable of which were had magnificent sails supported by neural spines off their back (dorsal) vertebrae.
Two famous examples include the carnivorous Dimetrodon and the herbivorous Edaphosaurus.
A. Edaphosaurus, B, Dimetrodon, and C. an early diapsid
Dimetrodon and Edaphosaurus are especially interesting since they lived at the same time and place but evolved their sails quite independently. Dimetrodon belongs to a group called the sphenacodonts that primitively lack a sail. Edaphosaurus belongs to a different group called the caseids that also primitively lack a sail.
A major question is, "why should two distantly related groups develop sails?"
To answer this we have to consider the possible function.
1. Sails were for display. In side view they make the animals look much larger, with little energetic cost.
2. The area of the sails would act as heat radiators or heat collectors if well-supplied with blood vessels.
These days, the latter explanation is favored. We don't know, however if their primary function was to warm the animals up or cool them down. Bramwell and Fellgett (1973) calculate that a 200 kg Dimetrodon would heat up from 26° C to 32° C in 205 minutes without a sail and only 80 minutes with a sail. In any case, it is likely that since a predator and a potential prey evolved them, this was a case of adaptive escalation, where it was important to gain or lose heat fast to keep as active as possible - to chase and be chased.
1. Differentiation of the tooth row into specialized regions. We can recognize teeth that we might call incisor-like, canine-like, and molar-like.
2. The fore limbs are relatively large and powerful compared to the hind limbs.
3. The rear bones of the lower jaw have become intimately involved with the ear region.This is quite different to the contemporaneous Sauropsida. In sauropsids -
1. There tends to be very little tendency for the tooth row to develop specialized regions (this is clearly a primitive feature retained from the ancestor of both synapsids and sauropsids).
2. The fore limbs tend to be relatively small and the hind limbs are often much larger (maybe a derived feature).
3. The rear bones in the lower jaw tend not to become associated with the ear (retention of a primitive feature).IV. In the late Permian more advanced synapsids, the therapsids, came to dominate the scene.
Therapsids are characterized by a reduction in both length and number of phalanges in the hand and foot. In addition there is a much greater specialization in the tooth row.
Carnivorous and herbivorous therapsids evolved rather quickly and the herbivores come to vastly outnumber their predators. By the end of the Permian, herbivorous therapsids include the numerically dominant dicynodonts, while the carnivorous forms are dominated by the cynodonts, which are especially mammal-like.
Importantly, one group of diapsids developed anteorbital and mandibular fenestrae. This group was the Archosauriformes. Although their Late Permian representatives are rare and small, they would be the group from which dinosaurs would evolve
By the middle Permian, ice vanished from the poles and a long hot-house era began that would last until the Cenozoic.
In the southern part of Pangea, so-called Gondwanaland, the flora was dominated by a group of seed plants, the most common genus of which was Glossopteris. Despite the fact that you could walk from southern Africa to North America, the flora of Gondwanaland remained distinct from that of the northern continents (Laurasia), where conifers dominated, until the beginning of the Triassic.
In the oceans, ammonites, which had evolved earlier in the Paleozoic, thrived, and huge reefs formed in the tropics.
V. As we have said, the largest mass extinction of all time brought an end to the Permian and to the Paleozoic as well.
Afterwards, within the Triassic, the reign of synapsid dominance came to an end and that of diapsid, particularly Archosauriformes, dominance began.
While we will leave the topic of the Triassic to the next lecture, we should consider what a typical early member of the Archosauriformes looked like. This will be our introduction to the anatomy of the Archosauriformes and will serve as a basis for understanding dinosaurian anatomy.
Euparkeria was a small, Early Triassic, carnivorous archosauriform from the Karoo basin of southern Africa. Its small fore limbs suggest it could run bipedally.
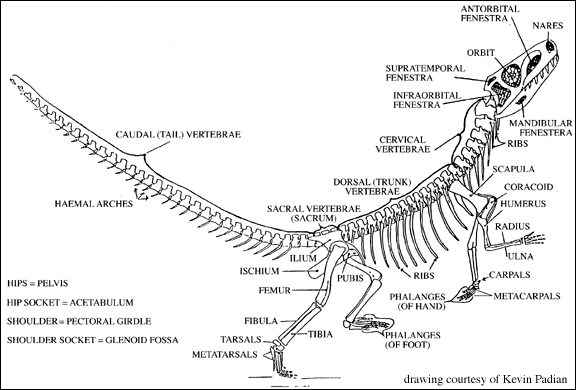
Skeleton of Euparkeria
 |
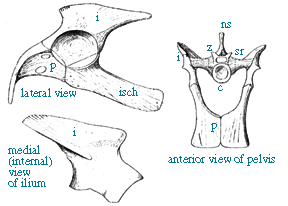 |
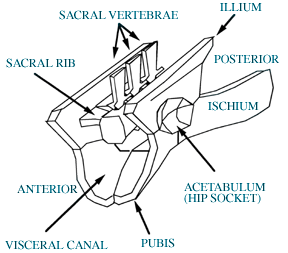
| Highly diagramatic view of archosauromorph pelvis comparable to that of Euparkeriain 3/4 view. | Reconstruction of the pelvis of Euparkeria capensis(from, Ewer, 1965). Abbreviations are: c, centrum of sacral vertebrae; i, ilium; isch, ischium; ns, neural spine, p, pubis; sr, sacral rib; z, zygopophysis; |
Ewer, R. F., 1965, The anatomy of the thecodont reptile Euparkeria capensis Broom. Philosophical Transactions of the Royal Society of London, Series B: Biological Sciences.248; 751, p. 379-435
http://www.ucmp.berkeley.edu/seedplants/pteridosperms/glossopterids.html (University of California Museum of Paleontology, Introduction to the Glossopteridales - home page at http://www.ucmp.berkeley.edu/index.html)
http://www.dinofish.com/ Web
site of the Coelacanth Rescue Mission, a project under the direction of
Jerome F. Hamlin to reduce
Coelacanth fatalities.
http://www.wcmc.org.uk/species/data/species_sheets/coelacan.htm (The World Conservation Monitoring Centre, Species Under Threat, Coelacanth - Latimeria chalumnae (Smith, 1939) - home page at http://www.wcmc.org.uk/