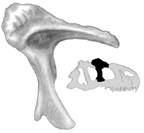
The Late Jurassic World, 150 Ma.
BACK TO DINOSAURS 2001 HOME PAGE
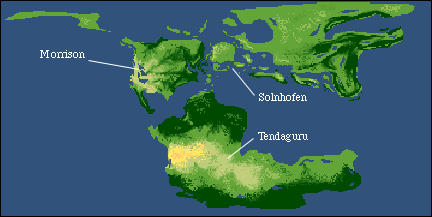
The Real Jurassic Park - Morrison and Tendaguru Formations
Although there are some excellent Middle Jurassic age assemblages of dinosaurs that are now coming to light, especially in China, we will be concentrating on Late Jurassic dinosaurs, simply because they are so well known. In this lecture we will examine the dinosaurs of the Morrison Formation of the western US and the Tendaguru Formation of Tanzania. In many respects, the two assemblages are quite similar. In the next lecture we will look at the fantastic preservation in the Solnhofen Limestone of Germany, which is about the same age as the Morrison and Tendaguru.
The Late Jurassic World, 150 Ma.
By Late Jurassic time the opening of the central Atlantic was well under way. However, land connections still existed between North America and Eurasia, and through Eurasia, the southern continents.

Outcrop of the Morrison Formation in Utah.
| What we know of Morrison vegetation itself, which is not much, is that it was dominated by conifer trees of the Cheirolepidiaceous kind (as earlier in the Jurassic) along with trees related to the modern Gingko, ferns, and horsetail rushes. An important point to remember is that flowering plants were not at all common, if they were present at all. Insect communities were getting quite modern in appearance. Termites are especially important insects because they eat (and hence respire) huge amounts of plant matter. Although fossil termite nests have been found in Triassic strata (Chinle), by the Late Jurassic some termites were building gigantic nests some 30 m in height. |
 Morrison termite nest, courtesy of Steven Hasiotis. |
The Morrison Formation is the most prolific source of Jurassic dinosaurs in North America. Thousands of bones of many hundreds of individuals have been found. The Morrison was one of the first units to produce bones during the Marsh and Cope wars and hence there is a long history of collection. Many fine museum exhibits of some of the most famous dinosaurs come from this formation.

Sauropod and Allosaurus bones excavated and left in situ at the display at Dinosaur National Monument near Vernal, Utah (Morrison Formation, Late Jurassic).
The Tendaguru strata differ from those of the Morrison in have marine interbeds. Little has been described about the climatic interpretation; however, continental reconstructions place the area in the southern hemisphere, but closer to the equator than the Morrison.
Overall the fauna of the Tendaguru is rather similar to the that of the Morrison. The most striking similarity is the presence of the giant sauropod Brachiosaurus in both formations. However, there are some notable differences, the most obvious of which is the absence of skeletons of large theropod dinosaurs comparable to the Morrison Allosaurus.
Cladogram of the Sauropodomorpha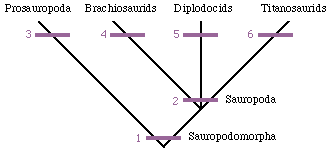 |
The most famous of the members of the Morrison dinosaur fauna are clearly the sauropods. A surprising diversity of sauropods have come from the Morrison and it is somewhat difficult to imagine how they effectively split up the available resources with such seemingly similar body plans. Like all sauropods, they have the shared derived character of having extra cervical vertebrae added to the neck at the expense of those of the dorsal series. The sauropods of the Morrison and Tendaguru fall into two broad groups (three by some counts). These are the diplodocids and the brachiosaurids. |
| Characters are - 1, small head and peg-like teeth (and elongated cervical vertebrae); 2, addition of extra cervical vertebrae to the neck; 3, apron-like pubis; 4, fore limbs as long as or longer than hind limbs, making the back slope posteriorly; 5, teeth restricted to front of mouth; 6, armor. | The diplodocids are characterized by having whip-like tails, low skulls, and teeth restricted to the front of the jaws. |
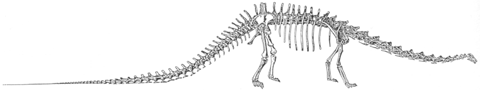
Diplodocus carnegii after Hatcher (1901).
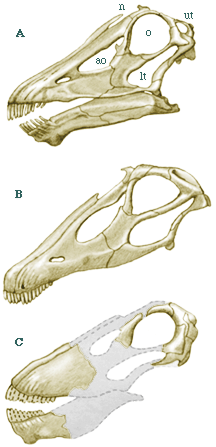 |
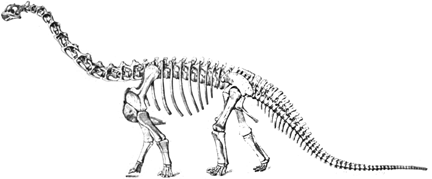
Diplodocus (A on left) is the genus from which the group derives its name. It was 27 m in length and an estimated weight of 10-11 metric tonnes (11-12 tons). Its skull is typical of diplodocids. Barosaurus is similar to Diplodocus except that its cervical vertebrae are 33 percent longer than in Diplodocus. The spectacular mounted skeleton in the Roosevelt Rotunda of the American Museum of Natural History is a Barosaurus. The American Musuem skelton is from the Morrison Formation but the genus has also been found in the Tendaguru. Seismosaurus is the longest known dinosaur according to David Gillette. It was between 39 and 52 m long (128-170 ft) and probably weighed in excess of 90 metric tonnes (100 tons). Apatosaurus (B on left) is probably the most famous dinosaur. Its more familiar name is Brontosaurus, although that name was given after the name Apatosaurus was applied. Apatosaurus was shorter but stockier than Diplodocus. For a long time the wrong head was associated with Apatosaurus. A Camarasaurus-like skull was made up and put on the Apatosaurus skeleton, which otherwise lacked a head. It turns out that a good candidate skull was found in the same quarry as Apatosaurus and fits the skeleton much better. It is a rather typical diplodocid skull, although distinctly different to Diplodocus itself. Dicraeosaurus (C on left) is a diplodocid from the Tendaguru that has an unusually short neck for this group. Abbreviations are - ao, antorbital fenestra; lt, lower temporal fenestra; n, nares; o, orbit, ut, upper temporal fenestra. Based on McIntosh (1990). |
| The Brachiosaurids are much more massively built than the diplodocids. Their shared derived character is having front legs as long as or longer than the hind legs. Thus these dinosaurs had a giraffe-like posture. Their namesake is of course Brachiosaurus. Brachiosaurus was first described from scrappy remains from the Morrison of Colorado and later from much more complete remains from the Tendaguru. As in other brachiosaurids, Brachiosaurus has nostrils placed high on the forehead and the front legs are considerably longer than the hind with the neck slanting upward following the upwardly sloping back. The Berlin mounted specimen of Brachiosaurus is 12 m (39 ft = 4 storeys) tall and 23 m (74 m long). | 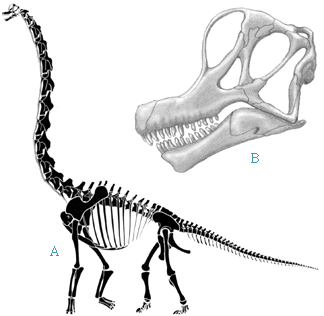 Brachiosaurus, redrawn after (A) Paul (1988) and (B) McIntosh (1990). |
Camarasaurus, after Gilmore (1925).
Camarasaurus (above) is a smallish sauropod at 18 m (59 ft). It is known from an excellent articulated skeleton. Camarasaurus does not have as proportionally long forelimbs as other brachiosaurids and the genus is often put in its own group, the camarasaurids, equal in taxonomic rank to the diplodocids, brachiosaurids, and titanosaurids.
Early on it was thought that sauropods, because of their obvious great bulk, had to be supported by water and that the tendency for the nostrils to be placed high on the skull indicated that they could breath underwater with just their nostrils protruding from the water.
What is wrong with this picture?It was also generally thought (and some still think) that sauropods ate soft water plants.The main problem is that the weight of water would crush the lungs.
But, there were no abundant soft water plants in the Jurassic! All large non-marine water plants suitable for eating are angiosperms (flowering plants).
| Another early idea was that sauropods had a lizard-like sprawling posture. However, their trunks are so deep that, if they had such a posture, their belly would drag on the ground. Footprints clearly show that the legs were carried directly under the belly, with no belly drag. They had a full upright posture and gait. On the right is a trackway of a sauropod of Early Cretaceous age in the bed of the Paluxy River, near Glen Rose, Texas. Abbreviations are - p, left pes impression; m, left manus impression. Note the lack of belly or tail drag and the relative narrowness of the trackway. |  |
There are several famous theropod dinosaurs from the Morrison Formation. The best known are Ceratosaurus nasicornis (a large late ceratosaur), Allosaurus fragilis (a large carnosaur), and Ornitholestes hermanni (a small basal coelurosaur). In contrast, the Tendaguru has produced only the small, fragmentary, possible ceratosaurian Elaphrosaurus.
Ceratosaurus nasicornis, adapted from Paul (1988).
Ceratosaurus nasicornis is a large theropod with a fused pelvis and a manus that still retains digit I. It is presently classified as a member of the Ceratosauria, and obviously gives its name to the group. Like many theropods, Ceratosaurus sports head ornaments, in this case a well developed, laterally flattened horn on the snout.
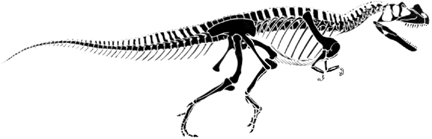
Allosaurus fragilis is a large theropod from the Morrison that is classified as a member of the Tetanurae because it has the groups' shared derived character of having lost digit IV and V on the manus. Like other tetanurians it also has a well developed "boot" or "foot" on the distal end of the pubis. Allosaurus is also a member of the Carnosauria, a supposedly-monophyletic group that has the shared derived character of having a large hole in the lacrimal bone of the skull. Originally, the Carnosauria was envisioned as being all of the large, carnivorous theropods. However, many theropods have been found to belong to other monophyletic groups that otherwise are comprised of modest- or small-sized theropods. Ceratosaurus is such an example, and Tyrannosaurus, now put within the Coelurosauria, is another. Furthermore, the supposed shared derived character of having a large cavity in the lacrimal bone is also seen in tyrannosaurs and the monophyly of the Carnosauria is in further doubt.
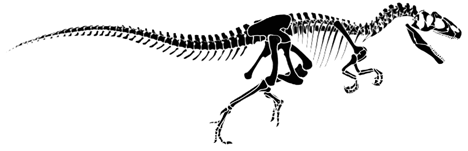
Allosaurus fragilis, redrawn from Paul (1988).
 Skull (cast) of Allosaurus in the Royal Ontario Museum, Toronto, Canada. |
The main shared derived character of the Carnosauria is a large cavity in the lacrimal bone of the skull. The lacrimal bone is located just at the antero-dorsal edge of the eye. The cavity may have held some sort of gland. Note also that the lacrimal forms a base for a horn as well - another example of theropod head ornamentation.
Lacrimal of Allosaurus (adapted from Anon., 1992). The "c" indicates the position of the cavity in the lacrimal. |
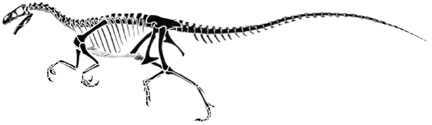
Ornitholestes, redrawn from Paul (1988).

Ornitholestes, a reconstructed skeleton (cast) at the Academy of Natural Sciences of Philadelphia.
Now we are in a position to look at the major groups of theropods examined so far. The summary cladogram, below, includes Ornitholestes, a group that includes Velociraptor (the Deinonychosauria), as well as birds plus Archaeopteryx and its relatives (the Avialae). The Deinonychosauria plus the Avialae comprise the Maniraptora, which we shall look at in the next lectures.
|
1. three-toed foot 2. digits IV and V lost on hand 3. long arms 4. semilunate carpal 5. fused pelvis 6. large hole in lacrimal bone in skull 7. ?no unique derived characters? 8. giant, hook-like claw on digit II of pes 9. flight feathers |
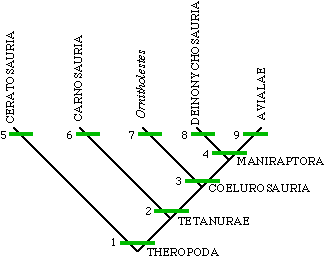 Cladogram of the Theropoda including Ornitholestes |
III. Ornithischia, Thyreophora
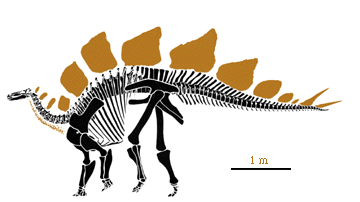
Stegosaurus stenops, adapted from Paul (1987).
Stegosaurus stenops was certainly the most spectacular ornithischian during Morrison time. The array of plates along the back with spikes limited to the tail place Stegosaurus within the Family Stegosauridae and within the Stegosauria. The shared derived character for the Stegosauria is armor as plates or spines on back.
|
The plates along the back of Stegosaurus have long been a puzzle, but recent work suggests that they may have served a thermoregulatory function, perhaps much like the sails of Dimetrodon or Edaphosaurus of the Permian. The plates themselves are riddled with blood vessel canals and hence it is difficult to see how they could not be of thermoregulatory significance. |
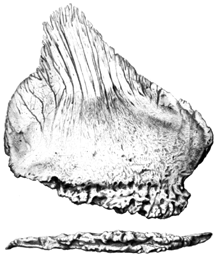 Dorsal plate of Stegosaurus, from Marsh (1896). |
Kentrosaurus aethiopicus is a stegosaurian from the Tendaguru. It differs from Stegosaurus in having spines on the back, shoulder, and hip as well as the tail. Stegosaurus-like plates are still present on the anterior part of the back and on the neck, however. Kentrosaurus is a member of the Family Kentrosauridae, the main shared derived character of which is the presence of spines on the back.
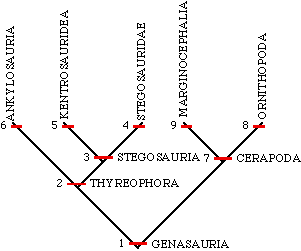
Below is a cladogram of the Genasauria that includes the Stegosauria and its members described above. Note that there are quite a range of other stegosaurs in the world and the group extends well into the Cretaceous.
|
1. cheeks 2. armor 3. armor as plates or spines on back 4. armor as plates 5. armor as spines 6. armor as covering or small plates over back 7. uneven enamel on teeth 8. jaw joint below tooth rows 9. bony shelf on back of head |
 |
One of the often-cited peculiarities of the Stegosauria is the very small size of their brain and the fact that there is a pronounced sacral swelling that has sometimes been called a "second brain". Below are natural casts of the brain cavity and the cavity of the sacral enlargement.
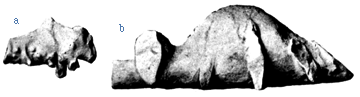
Endocast of brain (a) and natural cast of the sacral enlargement (b) of Kentrosaurus aethiopicus (from Hennig, 1915).
Many tetrapods have an enlarged part of their spinal cord in the sacrum. However, as is true for the endocast, the natural cast of the chamber to hold the nervous tissue is probably at least somewhat larger than the space occupied by the brain or sacral enlargement themselves. The remainder of the space was probably filled with connective tissue or even fat. Nonetheless, it is true that the brain is very tiny for such a large animal.
IV. Ornithischia, Ornithopoda
The best known cerapodans from the Morrison and Tendaguru are also early members of the Iguanodontidae. These are Camptosaurus dispar and Dryosaurus altus, respectively. To fully understand the position of these iguanodontians within the Ornithopoda, however we should look at the Early Cretaceous Hypsilophodon foxii from the Wealden Formation of the Isle of Wight, England.
Hypsilophodon was a small herbivore known from very complete and well-preserved material. The skeleton is shown below.
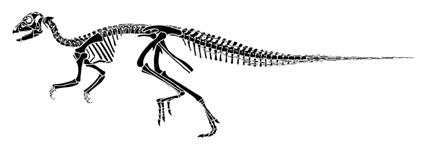
Hypsilophodon foxii, redrawn from Galton (1974).
The shared derived character for the Hypsilophodontidae, the family to which Hypsilophodon belongs, is the presence of a strong ridge on the side of the teeth. The Hypsilophodontidae share with the Iguanodontidae (and the Hadrosauria) a well developed anterior projection on the pubis. The usual ornithischian posterior process on the pubis, is of course, still present.
Remains of hypsilophodontids are known from the Morrison and include the little Othniellia rex, which is known from relatively fragmentary material.
The Iguanodontidae is characterized by the shared derived character of having a spike-like digit I on the manus. In Iguanodon itself this is a massive spike, but in Camptosaurus and Dryosaurus, the spike is rather small. Camptosaurus and Dryosaurus are shown below.
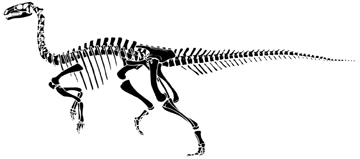
Camptosaurus dispar from the Morrison Formation (redrawn from Norman and Weishampel, 1990).
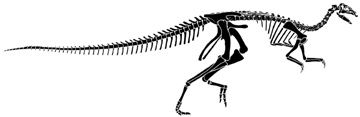
Dryosaurus altus from the Tendaguru Formation (redrawn from Sues and Norman, 1990).
Below is a cladogram depicting the relationships of the groups of Cerapoda we have covered so far.
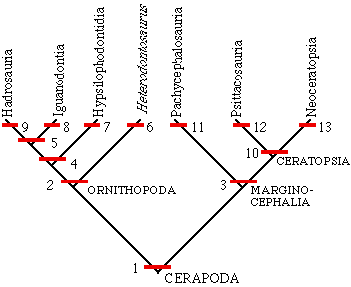 |
1. uneven enamel on teeth 2. jaw joint below level of tooth rows 3. bony shelf on back of head 4. prepubic process well developed 5. reduction in digits I and V on hand (and large nares) 6. elongate hand 7. strong ridge on side of teeth 8. spike-like digit I on hand (and very large nares) 9. duckbill like snout 10. rostral bone on snout 11. dome-like thickening on skull 12. reduction in manus 13. long frill |
In addition to the dinosaurs described above the Morrison, especially, has produced a wealth of other creatures, including turtles, crocodylomorphs (including crocodiles), true lizards and sphenodontians, fragmentary pterosaurs, and mammals.
Galton, P. M, 1974, The ornithischian dinosaur Hypsilophodon from the Wealden of the Isle of Wight. Bulletin of the British Museum (Natural History), Geology, v. 25, p. 1-52.
Gilmore, C. W., 1925, A nearly complete articulated skeleton of Camarasaurus, a saurischian dinosaur from the Dinosaur National Monument. Memoirs of the Carnegie Museum, v. 10, p. 347-384.
Hatcher, J. B., 1901, Diplodocus (Marsh): Its osteology, taxonomy, and probable habits, with a restoration of the skeleton. Memoirs of the Carnegie Museum, v. 1, p. 1-63.
Hennig, E., 1915, Kentrosaurus aethiopicus, der Stegosauridae des Tendaguru. Sitzungsberg. Ges. Naturforsch. Freunde Berlin 1915, p. 219-247.
McIntosh, J. S., 1990, Sauropoda. In Weishampel, D. B., Dodson, P., and Osmolska, H. (eds.), The Dinosauria, University of California Press, Berkeley, p. 345-401.
Marsh, O. C., 1896, Dinosaurs of North America. In Sixteenth Annual Report of the United States Geological Survey, pt. 1, p. 186, 196.
Norman, D. B. and Weishampel, D. B., 1990, Iguanodontidae and related Ornithopods. In Weishampel, D. B., Dodson, P., and Osmolska, H. (eds.), The Dinosauria, University of California Press, Berkeley, p. 510-533.
Paul, G. S., 1987, The science and art of restoring the life appearance of dinosaurs and their relatives. In Czerkas, S. J. and Olsen, E. C., (eds.), Dinosaurs, Past and Present, Vol. II, Natural History Museum of Los Angeles County, p. 5-49.
Paul, G. S., 1988, Predatory Dinosaurs. New York, New York Academy of Sciences, 464 p.
Sues, H.-D. and Norman, D. B., 1990, Hypsilophodontidae, Tenontosaurus" Dryosauridae. in Weishampel, D. B., Dodson, P., and Osmolska, H. (eds.), The Dinosauria, University of California Press, Berkeley, p. 498-509.